
Семена биология 6 класс: Строение семян — урок. Биология, 6 класс.
Тема урока: «Строение семян». 6-й класс
Тема урока: «Строение семян». 6-й класс- Ядренцева Ирина Эдуардовна, учитель биологии
Разделы: Биология
Класс: 6
УМК.
 Живой организм. 6
класс”/ Н.И. Сонин. – М.: Дрофа, 2013.
Живой организм. 6
класс”/ Н.И. Сонин. – М.: Дрофа, 2013.Оборудование.
Оптимальный вариант: семена фасоли и пшеницы, мультимедийный проектор, компьютер, электронное приложение к учебнику, электронный учебник для 6 класса линии “Навигатор” Сонина Н.И., электронные образовательные ресурсы по теме http://fcior.edu.ru/, презентация.
Минимум: фотографии и рисунки органов растений и животных.
Выдержка из программы основного общего образования. Биология. 5-9 классы. Авторы:
Место урока/
Урок №19 в разделе 1 “Строение живых организмов”.
Предыдущие уроки: “Органы цветкового растения. Цветок. Плод”.
Следующие уроки: “Основные системы органов животных”.
| Содержание урока | Характеристика видов
деятельности учащихся. |
| Строение семян Типы семян.
Строение семян однодольного и двудольного
растений Лабораторные и практические работы Строение семян |
Определяют понятия “зародыш”, “двудольные”, “однодольные”, “вегетативные органы”, “репродуктивные органы”. Анализируют типы семян. Узнают и запоминают строение семян двудольных и однодольных растений. |
| Предметные результаты обучения (для раздела) | Метапредметные результаты
обучения (для раздела) |
Учащиеся должны знать:
Учащиеся должны уметь:
|
Учащиеся должны уметь:
|
Цель урока: изучение строения семян
двудольных и однодольных растений.
Задачи урока.
1. Обеспечить достижение предметных результатов обучения.
Сформировать знания:
- о строении семян,
- однодольных растениях,
- двудольных растениях.
Сформировать умения:
- описывать внешнее строение семени,
- определять и называть части семени,
- выделять существенные признаки однодольных и двудольных растений,
- на основе строения семени растения определять принадлежность растения к классу однодольных или двудольных.
2. Обеспечить достижение метапредметных результатов обучения: создать условия (учебные ситуации) для развития коммуникативных, регулятивных и познавательных УУД.
3. Обеспечить достижение личностных результатов обучения:
- сформировать позитивное эмоциональное отношение обучающихся к изучаемым на уроке биологическим объектам,
- вызвать удивление уникальностью строения семян;
- сформировать понимание практической
значимости и ценности знаний о семенах для
выращивания культурных растений.

Формы работы: в парах, фронтальная, индивидуальная.
Структура и ход урока
Этап урока. (Приложение)
1. Организационный.2. Актуализация знаний.
3. Целеполагание.
4. Изучение нового материала.
5. Проверка понимания нового.
6. Подведение итогов.
7. Домашнее задание.
8. Рефлексия.
Приложение 1
Приложение 2
Приложение 3
Биология Строение семян. Изучение строение семян двудольных растений
Материалы к уроку
Конспект урока
Тема: Строение семян. Изучение строение семян двудольных растений.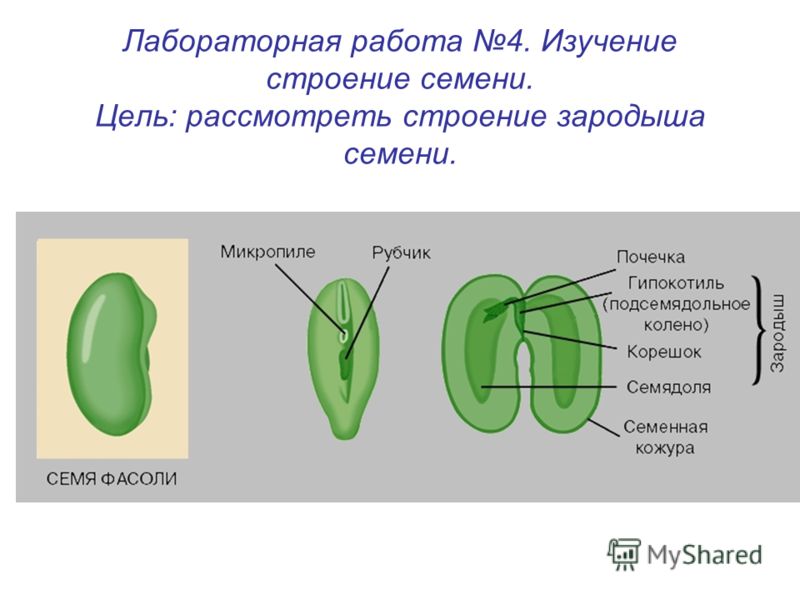
Ход урока
1. Организационный момент. Постановка целей урока
Здравствуйте, ребята! Я очень рада приветствовать Вас сегодня на уроке биологии.
Сегодня мы продолжим путешествие по удивительному царству растений, отделу покрытосеменных растений. Познакомимся с понятием двудольных растений.
2. Актуализация знаний
Но сначала я проверю, как вы усвоили материал домашнего задания.
1. Чем могут размножаться растения?
(Растения размножаются спорами, семенами, вегетативно).
2. Какие группы растений могут размножаться семенами?
(Семенами могут размножаться голосеменные и покрытосеменные растения).
3. Назовите отличие покрытосеменных растений от голосеменных растений?
(Отличие состоит в расположении семян. У голосеменных растений семена лежат открыто на поверхности шишек, у покрытосеменных семена находятся внутри плода).
4. Какие существуют растения по продолжительности жизни?
(По продолжительности жизни различают однолетние, двулетние и многолетние).
5. Как по-другому называют покрытосеменные растения?
(По – другому покрытосеменные называются цветковые, потому что они имеют органы семенного размножения — цветок).
6. Какие жизненные формы растений вам известны?
(деревья, кустарники, травы).
7. Назовите покрытосеменные растения, которые произрастают в нашей местности.
(тополь, подорожник, одуванчик).
8. Назовите самое высокое дерево в мире?
(Секвойя). Её высота – 115,5 метров. Рекорд установлен в 2006 году. В одной СЕКВОЙЕ индейцы прорубили отверстие – девятиметровый туннель.
9. Вспомните, как называется самая высокая в мире трава?
(бамбук). Каждый день молоденький БАМБУК вырастает на 50 см, но прибавляет он в росте не долго, всего 40 дней, а иначе дотянулся бы до облаков. Из стеблей этой травы делают удочки, лыжные палки, мебель, предметы быта. Очень красивый звук издают бамбуковые трубки.
10. Мякоть созревших плодов этого ДЕРЕВА варят, сушат, пекут в золе? (с
(ХЛЕБНОЕ ДЕРЕВО).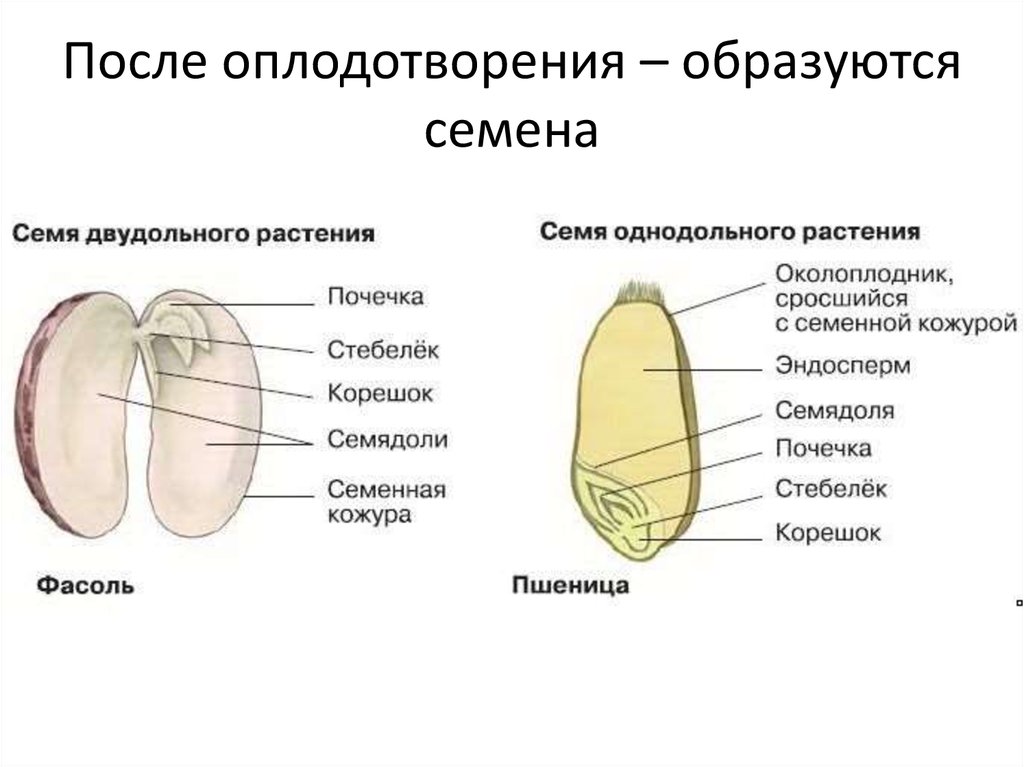
На нём растут овальные тяжёлые плоды, весом до 30 килограммов каждый. Из их мякоти местные жители пекут в золе лепёшки, похожие по вкусу на настоящий хлеб.
3.Изучение нового материала
Вот какие удивительные растения существуют на нашей планете, а как они появляются на свет, это нам и предстоит узнать сегодня на уроке.
На этом уроке мы начнем изучать семя. Что такое семя?
Семя – это генеративный орган, предназначенный для размножения и распространения семенных растений.
Это и есть зародыш будущего растения. В неблагоприятных для растения условиях этот зародыш может долгое время пребывать в состоянии покоя, то есть семя не будет прорастать. Поэтому мы можем хранить семена растений по несколько лет. Но когда мы посадим наши семена, они попадут в благоприятные условия и прорастут.
Семена растений бывают разные! Вспомните, как выглядят семена гороха и фасоли. Какого они размера? А какие семена поменьше? Ну а совсем маленькие? Семена некоторых растений, например, орхидеи Венерина башмачка, могут весить всего миллионные доли грамма.
А какой различной формы могут быть семена! Вспомните, какой формы семена яблока, груши?
А семена каких растений имеют форму шара? Некоторые семена имеют специальные крючки, как семена череды и лопуха.
Для чего им нужны эти крючочки?
Предположительные ответы учащихся
Различные выросты и крючочки им нужны, чтобы прикрепляться к животным и таким образом перемещаться на новые места обитания. А у некоторых растений есть на семенах пуховые выросты. У каких растений семена опушены? А для чего нужны эти выросты? Семена этих растений распространяются ветром. У семян клена и ясеня есть крылышки.
Для чего они нужны семенам?
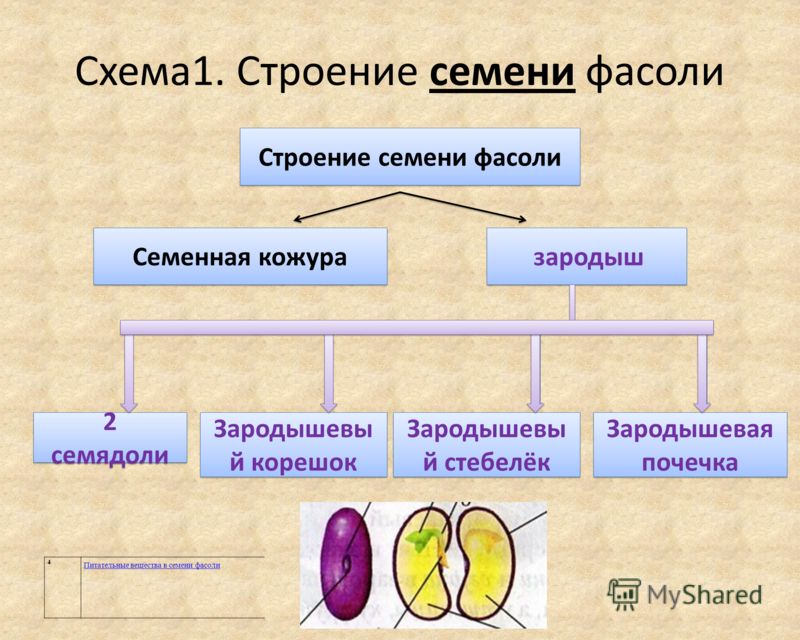
Давайте рассмотрим семя фасоли.
Лабораторная работа
“Строение семени фасоли”
Цель работы: изучить строение семени фасоли.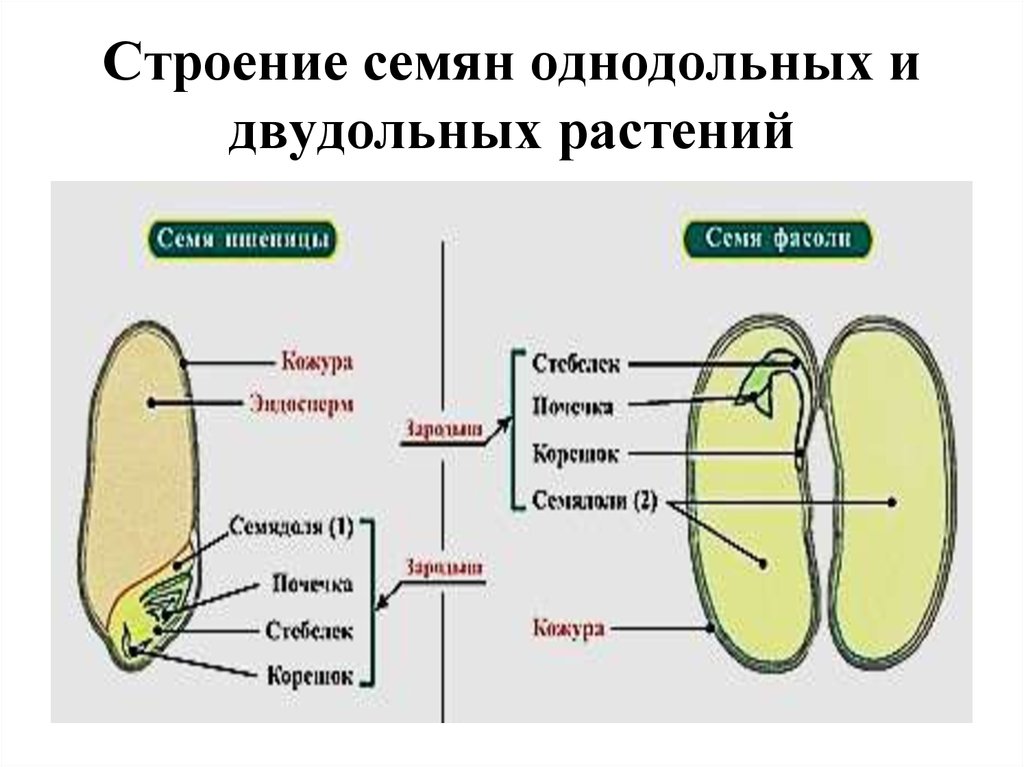
Материалы и оборудование: на каждую парту – 2 набухших зерна фасоли, 2 препаровальные иглы, 2 ручные лупы.
Ход работы:
1. Рассмотрите и устно опишите внешний вид семени фасоли (форма, поверхность, величина). Где расположен рубчик?
На брюшной стороне хорошо заметен след от семяножки, прикреплявшей семя к стенкам плода. Это рубчик, рядом — маленькое круглое отверстие — семявход.
Познакомимся со строением семени фасоли. Оно крупное, и все его части можно легко рассмотреть. Извлечем семя из плода, намочим его в воде и рассмотрим. Семя фасоли почковидное, уплощенное, снаружи покрыто толстой семенной кожурой.
Кожура — плотный, прочный наружный покров, белая или различно окрашенная (зависит от сорта).
Она надежно защищает семя от механического повреждения, высыхания, патогенных микроорганизмов и не дает ему прорасти до надежного и устойчивого установления благоприятных условий. Недаром мы поместили семена в мокрую тряпочку за 1,5-2 дня до урока, внимательно следили, чтобы она не высыхала, и содержали их в теплом месте. При мимолетном намокании вода внутрь не пройдет, т.к. сухая кожура плотно охватывает семя. У кожуры есть и другие функции, о них поговорим позже.
Недаром мы поместили семена в мокрую тряпочку за 1,5-2 дня до урока, внимательно следили, чтобы она не высыхала, и содержали их в теплом месте. При мимолетном намокании вода внутрь не пройдет, т.к. сухая кожура плотно охватывает семя. У кожуры есть и другие функции, о них поговорим позже.
2. Возьмите набухшее семя фасоли и отделите кожуру от зародыша.
Снимем семенную кожуру. С намоченного семени она снимается легко, а сухого удалить ее очень трудно.
С набухшего семени кожура снимается легко, и обнажаются семядоли — первые, зародышевые, листья, а значит, они являются частью зародыша. Семядоли толстые, мясистые, т.к. содержат много питательных веществ. Раздвигать семядоли начинаем со спинной стороны, медленно и осторожно. Если повезет, обе дольки останутся сидеть на маленьком побеге, но в любом случае легко увидеть места их прикрепления.
Зародышевый корешок уже приготовился выйти наружу. Осторожно снимаем кожуру.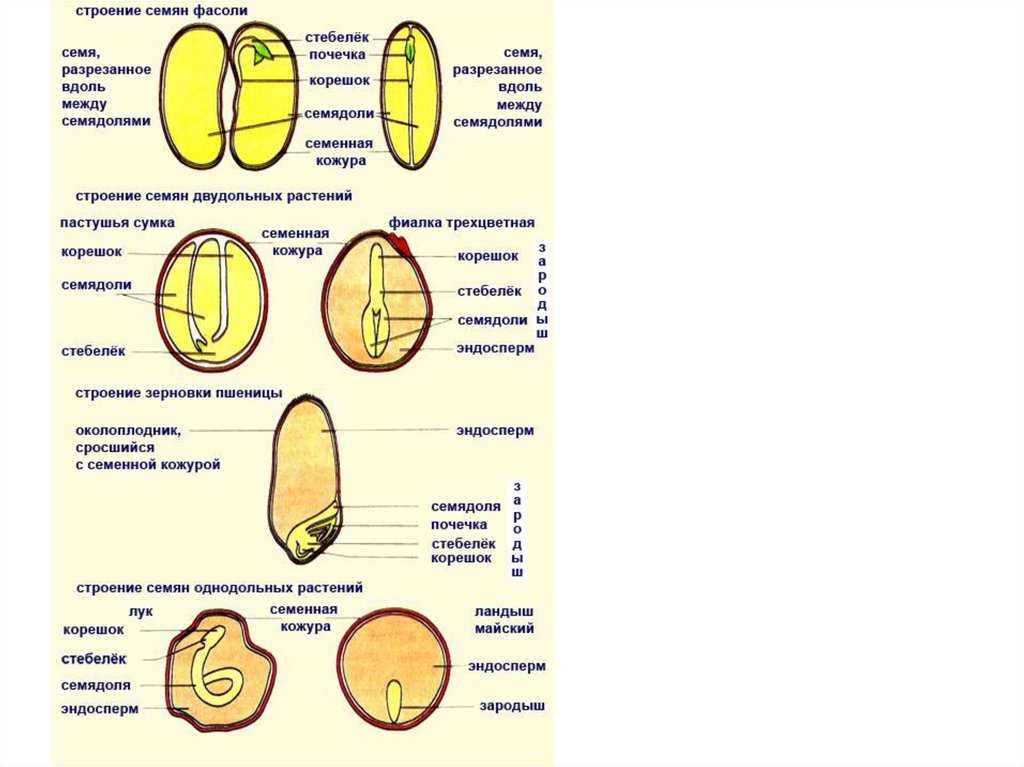
3. Прикрепите в тетради снятую кожуру и зародыш (или зарисуйте).
4. Возьмите целиком зародыш, рассмотрите его, найдите 2 семяпочки, корешок, стебелек, почечку.
5. Покажите основные части по схематическому рисунку. Какой вид имеет корешок, почечка, как их отличить? К какому органу зародыша прикрепляются семядоли?
Без видимой границы корешок плавно переходит в зародышевый стебель, на котором и сидят семядоли. В верхней части стебелек изгибается, унося внутрь семени от щели между семядолями зародышевую почку.
6. В тетради прикрепите (или зарисуйте) по отдельности 2 семядоли и остальные 3 части зародыша вместе. Подпишите их названия.
Какой орган зародыша является самым крупным? Почему семядоли толстые и большие? Каково строение зародыша? Для первого знакомства наиболее подходит фасоль. Для удобства назовем слегка выгнутую сторону семени спинной частью, вогнутую — брюшной.
Через него к зародышевой семяпочке еще в цветке проникают мужские половые клетки — пылинки, и тогда происходит оплодотворение. Он и теперь сослужит хорошую службу. Сжимаем набухшее семя — через семявход выступает капелька воды. Какой можно сделать вывод?
Он и теперь сослужит хорошую службу. Сжимаем набухшее семя — через семявход выступает капелька воды. Какой можно сделать вывод?
Через него в семя проникает вода!
— Где остановился корешок?
— Как раз напротив семявхода!
— Да, ему полагается первому выскочить наружу, закрепить семя в почве и начать добывать воду. С какими-то семенами это уже произошло.
— А где же эндосперм? Чем же будет питаться зародыш?
«Упитанный» вид семядолей наводит на мысль:
— А семядоли на что?
Дело в том, что у фасоли и других ее родственников (бобовых) семядоли оказались особенно трудолюбивыми и заботливыми няньками зародыша. Они заблаговременно перекачали все питательные вещества из эндосперма в себя. Окруженный такой заботой зародыш сформировал зачатки всех вегетативных органов будущего проростка, и при благоприятных условиях прорастание происходит очень быстро.
На этом их забота о зародыше не заканчивается.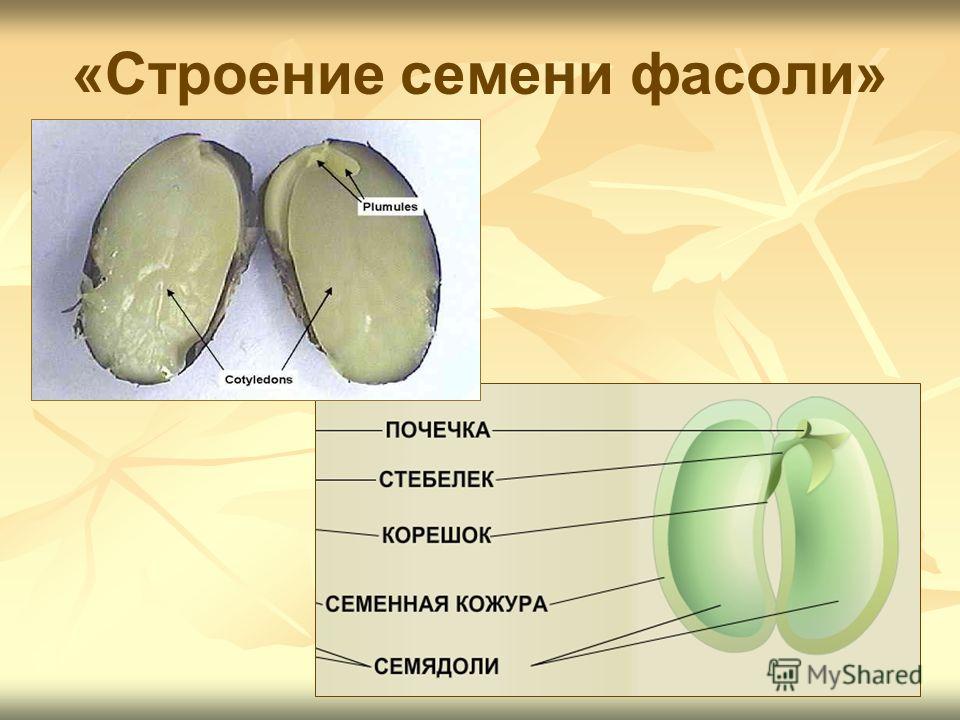
В плотных дольках, как в люльке, надежно укрылась нежная почка, и во время прорастания именно семядоли пробьют дорогу молодому побегу сквозь грунт, защитив почку от повреждения.
А молодые листики могут не спешить приступать к своим обязанностям — кормить растение продуктами фотосинтеза. И эту обязанность на первых порах возьмут на себя семядоли. Выйдя на поверхность, они заметно подрастут, позеленеют, и мы увидим на них многочисленные жилки проводящей системы. Они активно будут кормить молодой побег. А когда он войдет в силу, сморщатся, засохнут и отомрут.
Вспомним, какие еще растения выносят на поверхность свои крупные семядоли. Это огурцы, тыквы, кабачки.
А вот горох — нет. Его семядоли остаются под землей и поддерживают растущий побег лишь находящимися в них веществами. Не у всех растений семядоли заранее вбирают в себя питательные вещества из эндосперма. Некоторые частично или полностью откладывают эту работу на момент прорастания. Перекачивание питательных веществ — сложный биохимический процесс, подобный перевариванию пищи.
Перекачивание питательных веществ — сложный биохимический процесс, подобный перевариванию пищи.
Семядоли вырабатывают специальные вещества, растворяющие эндосперм, и превращают его в легко усваиваемую пищу для зародыша. Это особое «детское питание», как и полагается малышам! Позже мы узнаем, что эндосперм — это не простое хранилище питательных веществ, он тоже возникает в результате оплодотворения, отчего питательные вещества приобретают особую ценность. Эндосперм самым наилучшим образом способствует закреплению и развитию всех полезных свойств и качеств обоих родителей. Лиши семя эндосперма (как собственно эндосперма, так и семядолей) — и зародыш развиваться не будет, погибнет, и семя не даст потомства. Семядоли есть у всех семенных растений, но их количество, форма и время работы различны.
Таким образом, зародыш обладает теми же вегетативными органами, что и взрослое растение. У зародыша есть корень и побег. Зародышевый побег состоит из стебелька, двух зародышевых листьев (семядолей) и почечки.
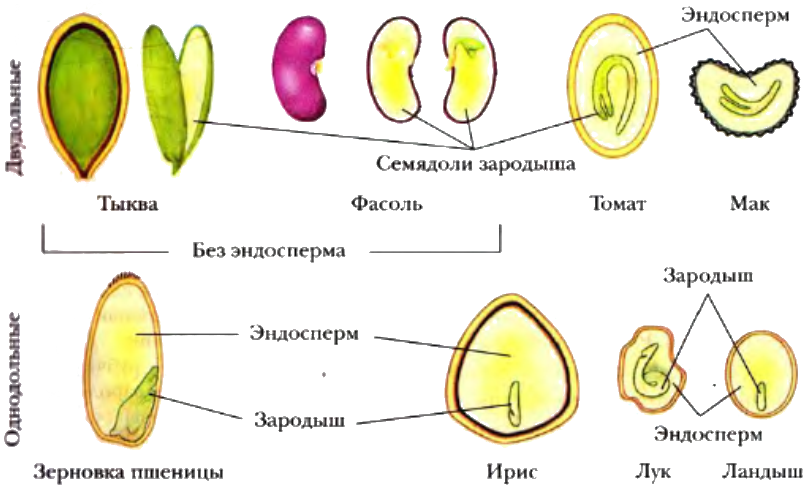
Растения, зародыш которых имеет две семядоли, относят к двудольным. Это — картофель, томаты, морковь, яблоня, дуб, огурцы и многие другие растения. Большинство двудольных растений имеет семена с эндоспермом.
4.Физкультурная минутка
|
Вновь у нас физкультминутка, |
И совсем не уставать. |
5.Закрепление знаний
Итак, что же мы узнали сегодня на уроке?
Мы вспомнили, как могут распространяться семена по планете, познакомились с понятием семя, узнали, какое строение оно имеет, познакомились с понятиями однодольное и двудольное растение, нашли маленькое растение в семени фасоли.
1.Цветок – орган семенного размножения имеют:
А. покрытосеменные растения
Б. голосеменные растения
В. папоротники
Г. лишайники
2. Семена развиваются внутри плода у:
А. голосеменных растений
Б. высших споровых растений
В. низших споровых растений
Г. покрытосеменных растений
3. Растения, которые выращивает человек, называются:
А. дикорастущие
Б. культурные
4. Господствующая группа растений на нашей планете:
А. голосеменные и покрытосеменные
Б. покрытосеменные
В. высшие споровые
Г. низшие споровые
5. Тополь, одуванчик, береза, дуб размножаются:
А. спорами
Б. делением клеток
В. семенами
Правильные ответы выделены жирным шрифтом
7. Рефлексия
«10 баллов»
Оценить по 10-бальной шкале работу на занятии с позиции:
„Я“ 0________10
„Мы“ 0________10
„Дело“ 0________10
8. Подведение итогов
Подведение итогов
Спасибо за внимание! Урок окончен.
Остались вопросы по теме? Наши репетиторы готовы помочь!
Подготовим к ЕГЭ, ОГЭ и другим экзаменам
Найдём слабые места по предмету и разберём ошибки
Повысим успеваемость по школьным предметам
Поможем подготовиться к поступлению в любой ВУЗ
Выбрать репетитора
6 класс. Биология. Строение семени. Плод — Строение семени. Плод
Комментарии преподавателяСемена покрытосеменных расположены внутри плодов. Они формируются из семязачатка в результате процесса двойного оплодотворения (один спермий оплодотворяет яйцеклетку, а другой – центральную клетку, с образованием запаса питательных веществ). Иногда околоплодник срастается с оболочкой семени, и семена практически неотличимы от плодов. Это происходит у злаковых (см. Рис. 1)
Рис. 1. Плоды злаковых растений
Семязачаток – женский гаметофит, проросший внутри женской споры, расположенной в спорангии.
Семя (см. Рис 2) состоит из семенной кожуры, зародыша и содержит запас питательных веществ.
Рис. 2. Строение семени
Семенная кожура может быть одно- или многослойной. Ее функция – защита зародыша от неблагоприятных воздействий окружающей среды.
Запас питательных веществ расположен в эндосперме, например, у злаковых растений (пшеница, овес), или в семядолях, как у бобовых (горох, фасоль).
В зародыше (см. Рис. 3) различают зародышевый корешок, стебелек, почечку и семядоли. Часто семядоли становятся первыми листьями растения.
Рис. 3. Строение зародыша
Однодольные растения – растения, семена которых состоят из 1 семядоли (злаковые, луковые, пальмовые и др.)
Двудольные растения – растения, семена которых состоят из 2 семядолей (картофель, томат, яблоня, горох и др.)
Семядоли
Не все зародыши двудольных растений имеют 2 семядоли, так же как и не все однодольные – одну.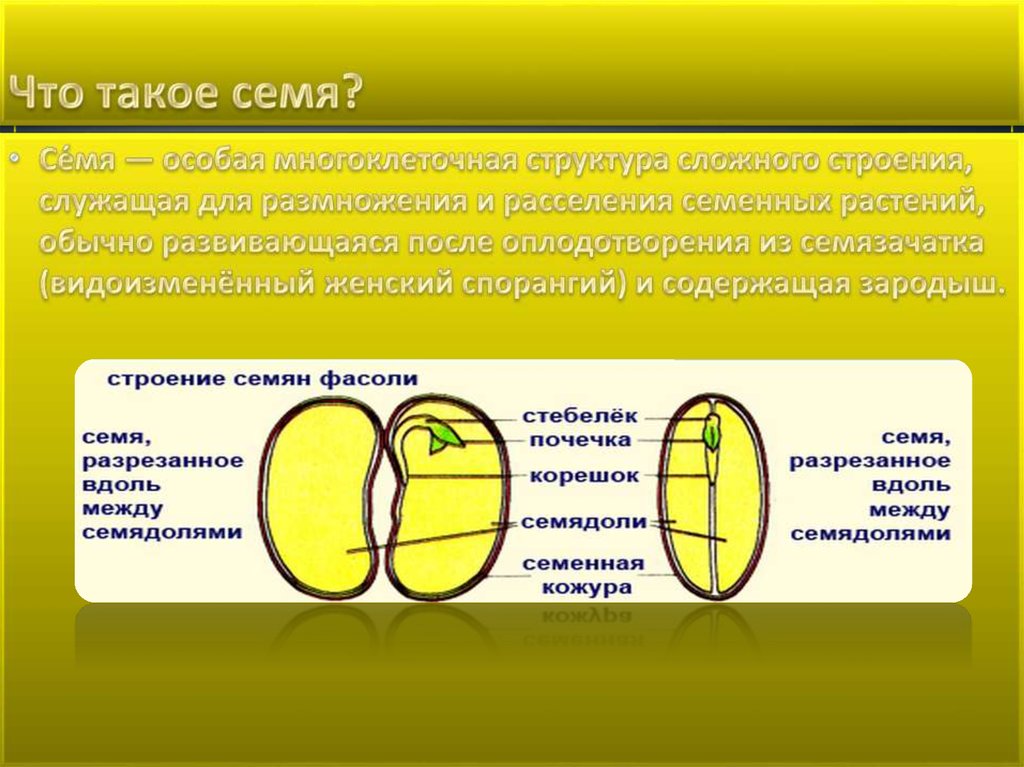 Так, в пылевидных семенах орхидей (класс однодольные) семядоли вообще отсутствуют.
Так, в пылевидных семенах орхидей (класс однодольные) семядоли вообще отсутствуют.
Голосеменные имеют многосемядольный зародыш, у некоторых хвойных насчитывается до 15 семядолей.
У миндаля зародыш разрастается настолько, что практически полностью вытесняет эндосперм. У стрелолиста, фасоли зрелое семя состоит только из зародыша и семенной кожуры. Весь запас питательных веществ расположен в клетках зародыша внутри семядолей.
Рубчик (см. Рис. 4) – место прикрепления семени к семяножке. Заметен на крупных семенах. С его помощью семя крепилось внутри завязи и получало питательные вещества.
Рис. 4. Рубчик
Микропиле (см. Рис. 5) – отверстие над рубчиком. Служит для проникновения в семя воздуха и воды.
Рис. 5. Микропиле
Рассмотрите сухие и набухшие, вымоченные в воде, семена фасоли. Сравните их размеры и форму. Снимите блестящую оболочку. Изучите зародыш: найдите семядоли, корешок, стебелек и почечку. Зарисуйте семя и подпишите название его частей. Определите часть семени, где расположены питательные вещества. Выясните, в каких частях семенидругие растения запасают питательные вещества. Рассмотрите строение семян яблони и тыквы.
Определите часть семени, где расположены питательные вещества. Выясните, в каких частях семенидругие растения запасают питательные вещества. Рассмотрите строение семян яблони и тыквы.
Семя пшеницы одето золотистым кожистым околоплодником. Они плотно сросся с семенем, их невозможно отделить. Поэтому правильнее говорить не о семенах пшеницы, а о плоде – зерновке.
Строение зерновки пшеницы
Рассмотрите форму и окраску зерновки пшеницы. Препаровальной иглой попробуйте снять часть околоплодника с набухшей и сухой зерновок. Объясните, когда она не снимается и почему. Рассмотрите под лупой разрезанную вдоль зерновку. Найдите эндосперм и зародыш (см. Рис. 6).
Рис. 6. Эндосперм и зародыш зерновки
Изучите строение зародыша. Зарисуйте зерновку пшеницы, подпишите названия основных частей. Попробуйте выяснить устройство семян других однодольных растений.
Семенадругих однодольных растений (лука, ландыша) имеют эндосперм, окружающий зародыш. У частухи (см. Рис. 7) созревшие семена эндосперма не имеют.
У частухи (см. Рис. 7) созревшие семена эндосперма не имеют.
Рис. 7. Частуха
Подкововидное семя состоит из кожуры и зародыша, в семядоле которого сосредоточены все запасы.
Механизмы распространения семян:
- Автохория – растение самостоятельно разбрасывает семена (недотрога, бешеный огурец)
- Анемохория – распространение семян с помощью ветра (клен, одуванчик, тополь)
- Анемогеохория – распространение семян с помощью перекатывания ветром по земле (перекотиполе)
- Гидрохория – распространение семян водой (кувшинки)
- Антропохория – распространение семян человеком (подорожник)
- Зоохория – распространение семян животными
- Эндозоохория – животные распространяют семена с экскрементами, поедая плоды (рябина)
- Эпизоохория – животные распространяют семена, прилипшие к их покровам (лопух)
- Орнитохория – распространение плодов птицами
- Мирмекохория – распространение плодов муравьями
Интересные факты
85% цветковых растений имеют семена с большим или маленьким эндоспермом.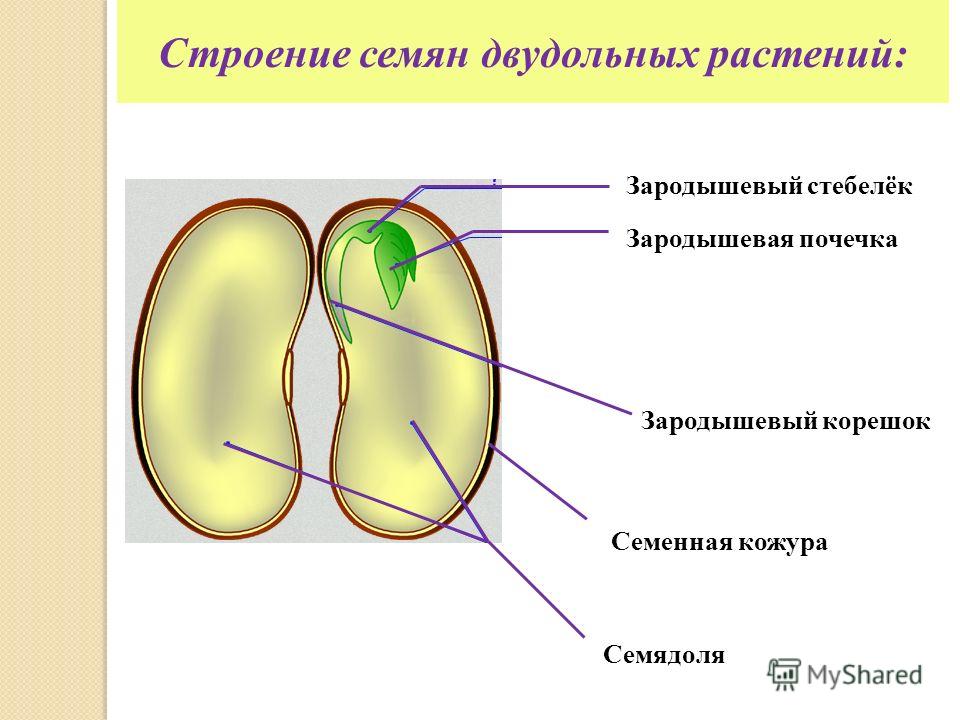
Самые маленькие размеры и массу имеют семена орхидных и некоторых паразитических растений.
Самые большие семена имеют растения сейшельской пальмы (см. Рис. 8). Их масса достигает 20 кг.
Рис. 8. Семена сейшельской пальмы
Прорастание – переход семени из состояния покоя в состояние вегетативного роста и формирование из него проростка. Период покоя семени является механизмом, обеспечивающим прорастание только при подходящих для этого условиях. Если во время прорастания семядоли выносятся на поверхность, то имеет место надземный тип прорастания (у фасоли, тыквы), а если остаются в почве – подземный (горох, пшеница).
источник конспекта — http://interneturok.ru/ru/school/biology/6-klass/bstroenie-pokrytosemennyh-rastenijb/stroenie-semeni-plod?seconds=0&chapter_id=738
источник видео — http://www.youtube.com/watch?v=edtH6v4f_X0
источник видео — http://www.youtube.com/watch?v=t4JetQt8REg
источник видео — http://www. youtube.com/watch?v=cHIWvZAU2DQ
youtube.com/watch?v=cHIWvZAU2DQ
источник видео — http://www.youtube.com/watch?v=HGoChiihLBo
источник видео — http://www.youtube.com/watch?v=hhtbmp7ikto
источник видео — http://www.youtube.com/watch?v=lieQBmCjx5c
источник видео — http://www.youtube.com/watch?v=_qnBdUlZTuA
источник видео — http://www.youtube.com/watch?v=g-IlbnaCgdM
источник презентации — http://ppt4web.ru/biologija/semja-vneshnee-i-vnutrennee-stroenie-semeni0.html
источник презентации — http://ppt4web.ru/biologija/plody4.html
Распространение плодов и семян 6 класс онлайн-подготовка на Ростелеком Лицей
Общие сведения
У многих растений распространяются не сами семена, а плоды. Эффективное распространение семян – залог выживания вида растения. Оно может происходить с помощью воды, животных, ветра, самораспространения.
Наблюдение: распространение семян
Понаблюдайте за распространением семян и плодов растений, растущих около вашего дома. Постарайтесь найти хотя бы по одному примеру для всех способов распространения. Приведите примеры растений, распространяющих семена подобным же образом.
Постарайтесь найти хотя бы по одному примеру для всех способов распространения. Приведите примеры растений, распространяющих семена подобным же образом.
Распространение ветром
Анемохория – распространение семян и плодов ветром.
Семена ивы, тополя, осины (см. Рис. 1) покрыты белым пухом и распространяются ветром на большие расстояния. Так же распространяются семена одуванчиков и ковыля.
Рис. 1. Семена ивы
Ветром могут распространяться только легкие сухие плоды. Для захвата ветра плоды имеют выросты, это плоды березы, ясеня, клена (см. Рис. 2). Сильный ветер может их сорвать с дерева только после полного опадания листьев, и так распространить.
Рис. 2. Плод клена
Плоды липы имеют характерные парусовидные листики (см. Рис. 3), что способствует их подхватыванию сильным ветром. Они, вращаясь, способны улетать от дерева на большие расстояния.
Рис. 3. Парусовидные листья плода липы
Распространение водой
Гидрохория – распространение семян и плодов растения водой.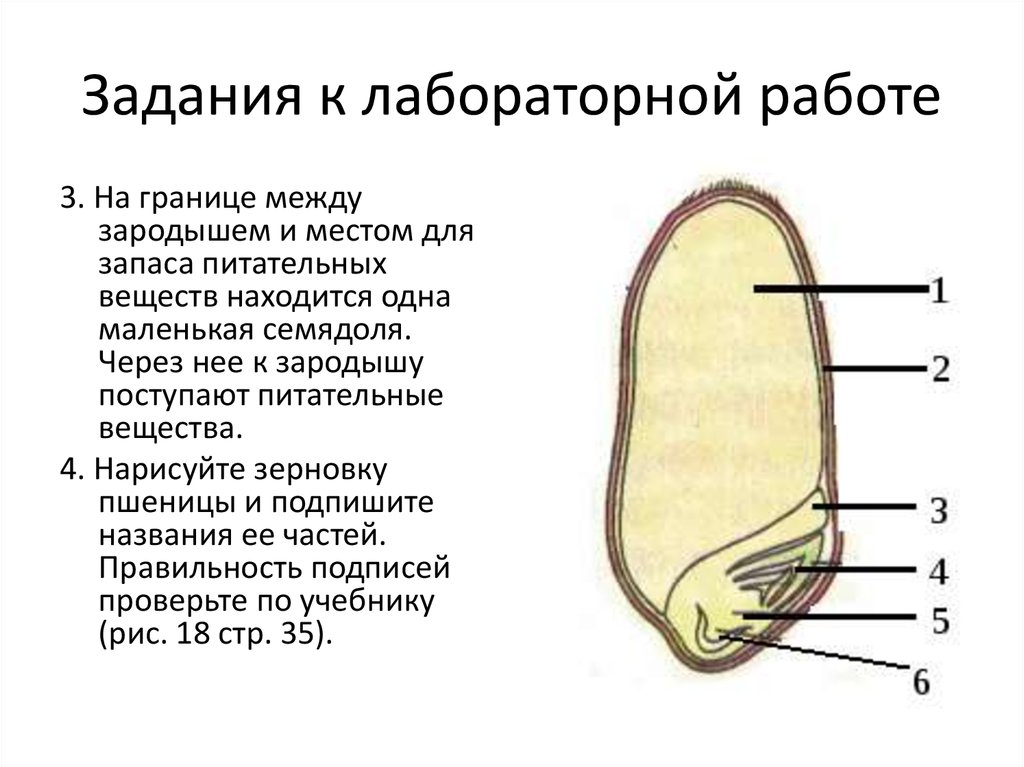
Анемогеохория
Некоторые степные растения образуют ветвистый, тонкий у основания стебель, который осенью, по мере созревания плодов, отмирает. Ветер отламывает растение у корня и перекатывает его. Такие растения называют перекатиполе. Примеры растений: качим, кермек.
Анемогеохория – способ распространения семян перекатыванием всего растения по земле ветром.
Плоды ольхи (см. Рис. 4), попадая в воду, не тонут и разносятся течением на мелкие расстояния.
Рис. 4. Плоды ольхи
Плоды кокосовой пальмы морскими течениями переносятся с одного острова на другой. Такие плоды устойчивы к соленой воде. Так, плоды сейшельской пальмы не тонут в воде, но гибнут в морской воде.
Распространение семян саморазбрасыванием
Автохория – распространение семян растений саморазбрасыванием (см. Рис. 5). Так, при задевании плода растения недотрога его створки резко скручиваются, разбрасывая семена. Это характерно для гороха, фасоли, других бобовых.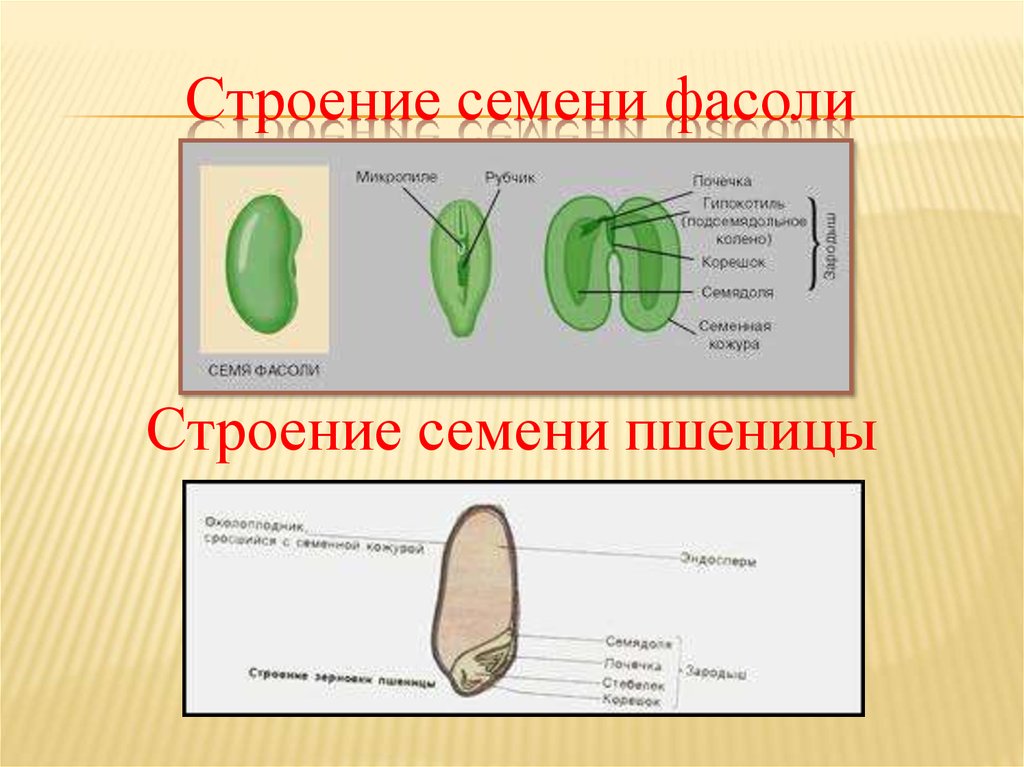 Семена разбрасываются на не очень большое расстояние.
Семена разбрасываются на не очень большое расстояние.
Рис. 5. Автохория фиалки
Распространение с помощью животных
Зоохория – распространение семян и плодов растений с помощью животных.
Бешеный огурец
На сухих склонах и морских побережьях можно встретить сорное растение бешеный огурец. После созревания семян в его плодах скапливается слизь, которая вместе с семенами может выстреливаться из плода. Такие семена могут прилипнуть к человеку или животному, задевшему плод.
Плоды лопуха (см. Рис. 6) или череды снабжены острыми зубчиками или крючками. С их помощью плоды прицепляются к шерсти животных или одежде людей, которые разносят их на большие расстояния.
Рис. 6. Плоды лопуха
Сочные плоды брусники (см. Рис. 7), черники, бузины и др. поедаются животными, в основном, птицами. Их семена защищены твердой оболочкой и не перевариваются. Поэтому выбрасываются вместе с пометом.
Орнитохория – распространение семян птицами.
Рис. 7. Плоды брусники
Желуди, каштаны (см. Рис. 8) падают прямо под родительское растение. Как правило, такие плоды питательные, и многие животные делают из них запасы корма на зиму. Часто животное гибнет или не способно найти спрятанные запасы. В этом случае растение прорастает.
Рис. 8. Плод каштана
Распространение с помощью человека
Антропохория – распространение семян растений с помощью человека.
При перевозке различных грузов семена и плоды растений могут прилипнуть к тюкам или мешкам. При их разгрузке семена могут отпадать и прорастать. Также семена могут прилипать к транспорту.
Перенос семян может происходить между континентами. Так, из Европы в Америку был завезен подорожник. Из Америки в Европу были завезены амброзия (см. Рис. 9) и ромашка пахучая.
Рис. 9. Амброзия
У одного вида растений могут сочетаться различные виды распространения плодов и семян. Так, семена фиалки, помимо саморазбрасывания, распространяются муравьями (имеют питательные придатки, привлекающие муравьев).
Плоды вяза распространяются ветром и водой.
Мирмекохория – распространение семян муравьями.
Список литературы
- Биология. Бактерии, грибы, растения. 6 кл.: учеб. для общеобразоват. учреждений / В.В. Пасечник. – 14-е изд., стереотип. – М.: Дрофа, 2011. – 304 с.: ил.
- Тихонова Е.Т., Романова Н.И. Биология, 6. – М.: Русское слово.
- Исаева Т.А., Романова Н.И. Биология, 6. – М.: Русское слово.
Дополнительные рекомендованные ссылки на ресурсы сети Интернет
- Biolicey2vrn.ucoz.ru (Источник).
- Cosmoschool.ru (Источник).
- School.xvatit.com (Источник).
Домашнее задание
- Биология. Бактерии, грибы, растения. 6 кл.: учеб. для общеобразоват. учреждений / В.В. Пасечник. – 14-е изд., стереотип. – М.: Дрофа, 2011. – 304 с.: ил. – с. 151, задания и вопросы 1, 5 (Источник).
- Какие приспособления имеют семена для распространения ветром?
- Что такое зоохория? Для каких растений она характерна?
- * Существует ли наиболее выгодный способ распространения семян? Какой? Аргументируйте как отрицательный, так и положительный ответы.

Презентация по биологии 6 класс Строение семени доклад, проект
Давайте вспомним!
Какие группы растений являются высшими, а какие низшими?
Почему водоросли относят к низшим растениям?
Назовите представителей одноклеточных зеленых водорослей
(хламидомонада, хлорелла).
Многоклеточные водоросли
(улотрикс, спирогира).
Как называют тело многоклеточных водорослей?
Что такое ризоиды и какова их роль?
А какие органы есть у покрытосеменных растений?
На какие две группы делятся эти органы?
Актуализация знаний
Какие органы растений вы знаете?
К вегетативным относятся …
К генеративным относятся…
Строение семян покрытосеменных растений
Семя – орган полового размножения и расселения растений.
Где находится семя?
Сколько их может быть в плодах?
Что такое семя?
Лабораторная работа
«Строение семени»
“Коробочка запечатана,
А в ней растенье спрятано.
С ветками листочками,
С белыми цветочками”
Чтоб найти нам ответ,
Мы исследуем объект.
Чем покрыто семя?
Строение семени
Кожура – плотный покров семени.
Главная функция: зашита от повреждений и высыхания, проникновения болезнетворных организмов и от преждевременного прорастания.
Строение семени
Рубчик – след от места прикрепления семени к стенке плода.
Семявход – маленькое отверстие, которое находится рядом с рубчиком, через которое в семя проникает вода, после чего семя набухает и прорастает.
Зародыш – это миниатюрный спорофит (бесполое поколение у растений, представленное диплоидной фазой и развивающееся из зиготы). Зародыш обычно образуется из оплодотворенной яйцеклетки и представляет собой зачаток новой особи.
Зародыш обычно образуется из оплодотворенной яйцеклетки и представляет собой зачаток новой особи.
Зигота — клетка, образующаяся в результате оплодотворения Из зиготы образуется новая особь (животное или растение)
Строение семени
Эндосперм – запасающая ткань семени.
Зародыш нового растения в семени имеет две хорошо различимые части: зародышевый побег и зародышевый корень.
Цветковые растения, имеющие зародыш семени с одной семядолей, называются однодольными, а с двумя – двудольными.
Совокупность всех корней растения называют корневой системой.
Многообразие семян
Семена одуванчика
Семена льна
Семена арбуза
Семена томатов
Семена фасоли
Строение семени
Строение семени
А – кукурузы;
Б – фасоли.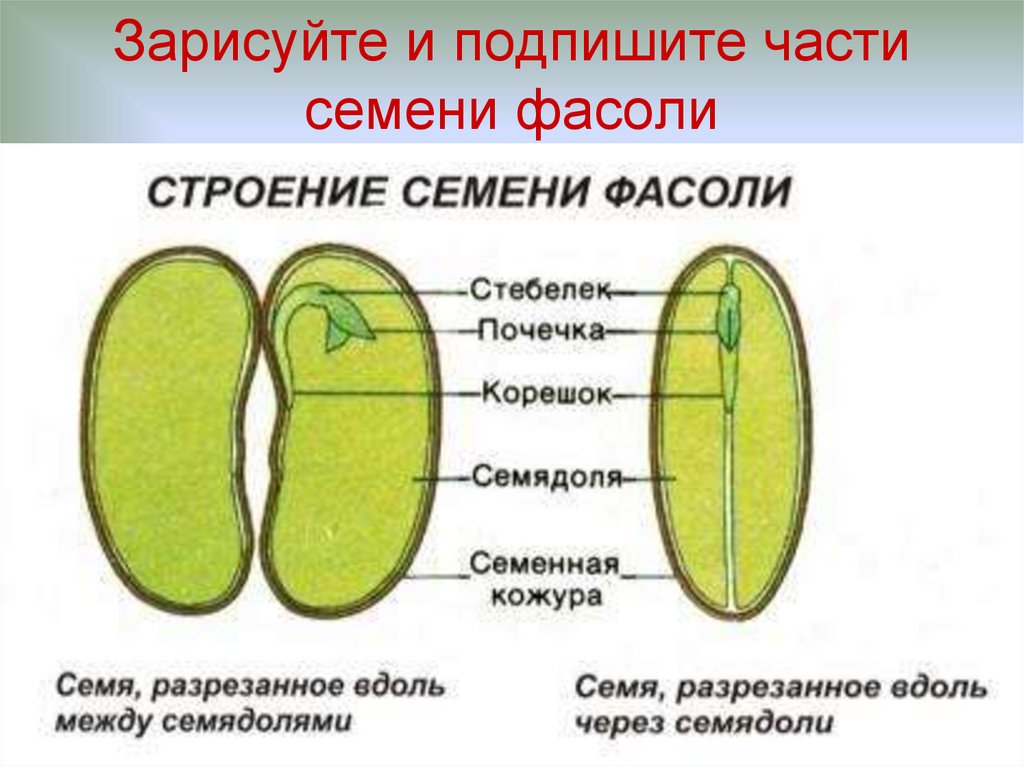
Строение семян кукурузы (А) и фасоли (Б): 1 — кожура семени; 2 — семявход; 3 — рубчик; 4 — зародыш; 5 — зародышевая почка; 6 — покровы плода зерновки; 7 — зародышевый корень; 8 — зародышевый стебель; 9 — эндосперм; 10 — семядоли
Вывод по лабораторной работе:
Условия прорастания семян
Прорастание – это переход семян из состояния покоя к росту зародыша и развитию из него проростка.
Что необходимо для прорастания семян?
Кислород;
Запасные питательные вещества;
Вода;
Температурные условия;
Влияние света.
Значение семян
Для чего нужны семена?
Какой околоплодник у этих плодов?
Яркий и сочный околоплодник..
Может такой околоплодник защитить семя? Нет.
Своим вкусом и цветом он привлекает животных.
Какую пользу из этого извлекают растения?
Животные, поедая плоды, распространяют семена растений
Вставьте пропущенные слова
Зародыш семени однодольного растения состоит из … …, … … и … … .
Зародыш семени однодольного растения состоит из зародышевого корешка , зародышевого побега и одной семядоли.
Двудольными называют растения, в семени которых…
Двудольными называют растения, в семени которых две семядоли.
Домашнее задание
Параграфы 10-12 – читать, учить по тетради;
Параграф 12 «Значение семян» – вопросы 1, 3 – пис.
Скачать презентацию
Презентация по биологии на тему строение семян(6 класс) доклад, проект
Строение семян однодольных и двудольных растений.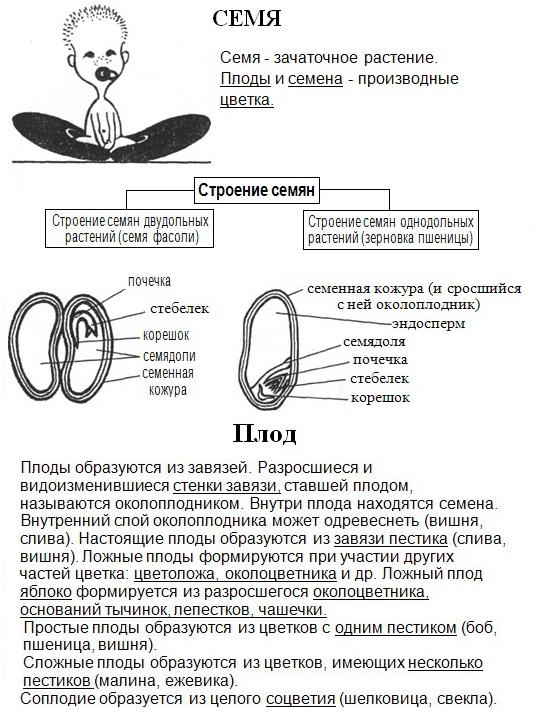 Условия прорастания семян.
Условия прорастания семян.
Выполнила: учитель биологии Aлиева Лейли
г.Имишл
Многообразие семян
Семена гороха
Семена фасоли
Семена редиса
Семена моркови
Плоды и семена
рожкового дерева
Семена рожкового дерева Вес одного семени — 200миллиграммов
Плоды рожкового дерева
Многообразие семян
Семя
Семя — особая многоклеточная структура сложного строения, служащая для размножения и расселения семенных растений, обычно развивающаяся после оплодотворения из семязачатка (видоизменённый женский спорангий) и содержащая зародыш.
семя клёна белого в разрезе
Семена бывают разных размеров: очень большие и микроскопически маленькие, и зарождаются они в разном количестве. Образование плода происходит с развитием семени.
Образование плода происходит с развитием семени.
Семена и строение семени
Семенная кожура́ — структура, снаружи покрывающая и защищающая зародыш в семени от перенасыщения влагой или пересыхания.
Семенная кожура
Семенная кожура манго (Mangifera indica)
Эндосперм — ткань, содержащаяся внутри семени, обычно окружающая зародыш и снабжающая его питательными веществами в ходе развития. У цветковых эндосперм обычно образуется в ходе двойного оплодотворения в результате слияния центральной клетки (центрального ядра) зародышевого мешка с одним из спермиев. У многих цветковых клетки эндосперма триплоидны. У кувшинки эндосперм образуется при слиянии спермия с гаплоидной клеткой зародышевого мешка, так что его ядра диплоидны. У многих цветковых ядра эндосперма имеют набор хромосом более чем 3n (до 15 n).
Эндосперм
Зародыш
Под кожурой находится зародыш — маленькое будущее растение.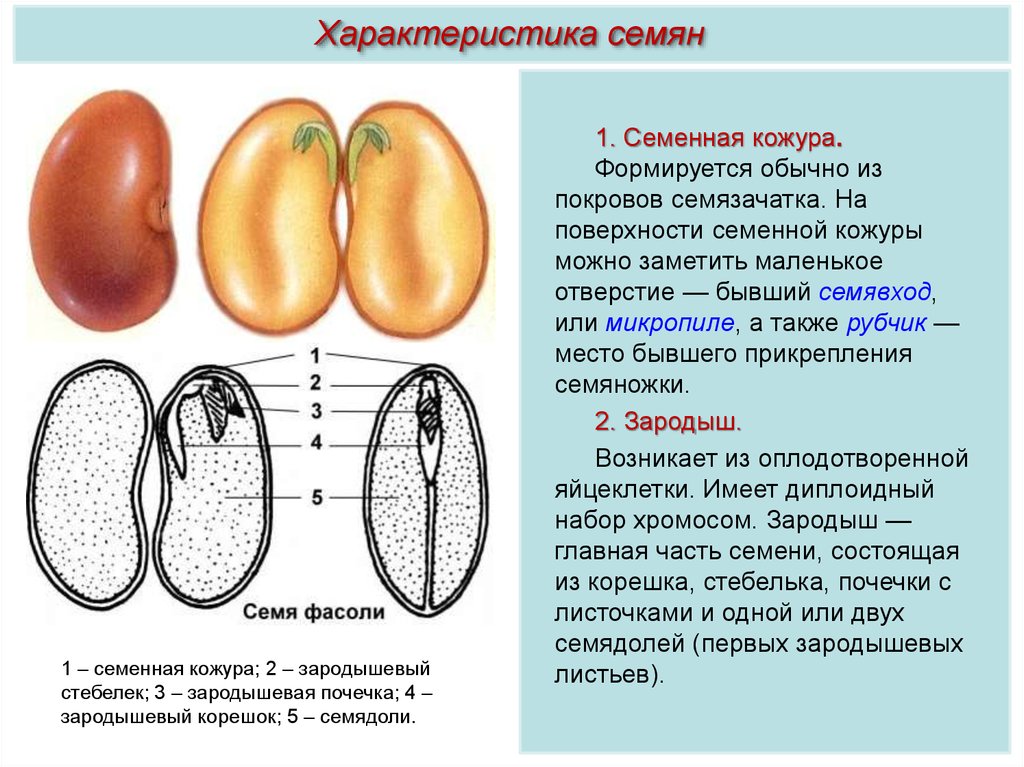 Зародыш у многих цветковых состоит из зародышевого корешка, зародышевого стебелька, зародышевой почечки и семядолей. У других групп (например, у подавляющего большинства орхидных) зародыш до прорастания семени не имеет дифференцированных органов.
Зародыш у многих цветковых состоит из зародышевого корешка, зародышевого стебелька, зародышевой почечки и семядолей. У других групп (например, у подавляющего большинства орхидных) зародыш до прорастания семени не имеет дифференцированных органов.
Прорастание семени гороха
семядоли
Зародышевый корень
Зародышевый стебель
Зародышевая почка
10*
Запас питательных веществ находится в семядолях
Минимальные размеры имеют семена орхидных и заразиховых; их масса составляет 0,001—0,003 мг. Максимального размера достигают семена сейшельской пальмы Lodoicea maldivica (масса около 20 кг).
Размер семян
Плод сейшельской пальмы
Семена орхидеи
Химический состав семян
Основная особенность химического состава зрелых семян — очень низкое содержание воды, обычно всего 10—15 % (по разным источникам, от 5 до 20 %).
Химический состав семян сильно зависит от условий созревания и от сорта растения.
Всхожесть семян — это их способность давать за определённый срок нормальные проростки (в лаборатории) или всходы (в полевых условиях). Всхожесть сильно зависит от условий проращивания и от условий хранения семян. Обычно всхожесть выражают в процентах (это процент семян, которые дали всходы, от общего числа семян).
При длительном хранении семян со временем их всхожесть падает. Семена некоторых растений теряют всхожесть уже через 2—3 недели (например, семена большинства видов ив полностью теряют всхожесть при температуре 18—20 °C в течение месяца). Всхожесть семян большинства культурных растений заметно снижается через 2—3 года. Семена лотоса в торфе сохраняют всхожесть не менее 250 лет (по некоторым данным, более тысячи лет). Сохранившиеся в вечной мерзлоте семена люпина арктического удалось прорастить через 10—12 тыс. лет.
Всхожесть семян
Условия, необходимые для прорастания семян
Текст слайда
Семя – зачаток нового растения
лист
стебель
корень
кожура семени
семядоли
стебелек
почечка
корешок
это переход семян из состояния покоя к росту зародыша и развитию из него проростка
Прорастание
Потребность в воздухе
и воде у разных семян
Зародыш семени может потреблять питательные вещества только в растворённом виде.
При прорастании семян зародыш интенсивно дышит, требуется постоянный приток кислорода.
Потребность в тепле
у разных семян
Для прорастания необходимо разное количество тепла. Растения, семенам которых при прорастании требуется высокая температура, называют теплолюбивыми, а прорастающие при низких температурах, называют холодостойкими.
Семена растений прорастают при положительной температуре. Температура начала прорастания широко варьирует у растений разных таксономических групп и географических регионов. В среднем семена растений полярных и умеренных широт прорастают при более низкой температуре, чем семена субтропических и тропических видов. Различаются также оптимальная температура прорастания, при которой наблюдается наибольшая всхожесть и максимальная.
Семена некоторых растений выдерживают периоды кратковременного воздействия высокой температуры во время лесных пожаров, после которых создаются благоприятные условия для прорастания выживших семян. Кроме того, огонь способствует вскрыванию плодов некоторых видов растений, обладающих устойчивостью к воздействию огня. Так, только после пожаров вскрываются «поздние» шишки сосны скрученной, шишки секвойядендрона и др., плоды некоторых видов рода банксия.
Кроме того, огонь способствует вскрыванию плодов некоторых видов растений, обладающих устойчивостью к воздействию огня. Так, только после пожаров вскрываются «поздние» шишки сосны скрученной, шишки секвойядендрона и др., плоды некоторых видов рода банксия.
Условия прорастания семян
Температура
Условия прорастания семян
Глубина заделки семян
различных растений
1 этап – Поглощение воды.
2 этап – Набухание семян.
3 этап – Увеличение размеров. Деление клеток.
4 этап – Появление корешка.
5 этап – Появление зародышевого побега.
Этапы прорастания семян
Типы прорастания семян
Прорастание однодольных
и двудольных семян
однодольные
двудольные
Саморазбрасывание семян (автохория)
Семена многих растений падают на землю рядом с материнским растением после вскрытия плодов. Иногда при вскрытии плодов семена с силой выбрасываются, разлетаясь на некоторое расстояние. Саморазбрасывание семян характерно для таких растений, как недотрога мелкоцветковая, кислица обыкновенная.
Иногда при вскрытии плодов семена с силой выбрасываются, разлетаясь на некоторое расстояние. Саморазбрасывание семян характерно для таких растений, как недотрога мелкоцветковая, кислица обыкновенная.
Распространение семян
недотрога мелкоцветковая
кислица обыкновенная
Семена многих растений распространяются ветром (анемохория). Это, например, семена сосны обыкновенной, снабжённые крылышком, семена растений родов Тополь и Ива, покрытые волосками («тополиный пух»), мелкие пылевидные семена орхидных.
Распространение ветром
Распространение водой (гидрохория)
Водой распространяются плоды и семена не только водных, но и некоторых наземных растений. Ольха часто растет по берегам рек; ее плоды, попадая в воду, не тонут. Течение уносит их далеко от материнских растений. Плоды кокосовой пальмы с одного острова на другой переносятся морским течением.
Распространение с помощью животных
Распространение с помощью животных — зоохория. Семена растений могут распространяться животными на теле (обычно вместе с плодами), при прохождении через кишечный тракт и при растаскивании с потерей семян.
На теле разносят семена и односемянные плоды обычно птицы и млекопитающие. Так, млекопитающие могут разносить на шерсти плодики гравилата, череды, репешка и многих других растений, обладающих крючками, волосками и прицепками. Также на теле птиц и млекопитающих могут распространяться клейкие семена омелы, кувшинки и др.
Через кишечники птиц и млекопитающих после поедания ими плодов проходят, не теряя всхожести, семена таких растений, как бересклет бородавчатый, боярышник, малина и многих других.
Делая запасы в кладовых, белки, бурундуки, сойки и кедровки теряют часть семян или не находят часть кладовых, способствуя распространению семян сосны сибирской и дуба.
Гравилат
Многие организмы (от грибов и бактерий до птиц и млекопитающих) питаются в значительной степени, а иногда и исключительно семенами. Семена составляют основу пищи таких животных, как некоторые насекомые и их личинки (например, муравьи-жнецы), зерноядные птицы, грызуны (бурундуки, белки, хомяки и др.).
Основу рациона человека со времён возникновения земледелия в большинстве регионов мира также составляют семена, в первую очередь, культурных злаков (пшеницы, риса, кукурузы и др.). Главное питательное вещество, с которым человечество получает наибольшее число калорий, — крахмал, содержащийся в семенах злаков. Важным источником белков для человечества служат также семена бобовых растений — сои, фасоли и др. Семена являются основным источником растительных масел, которые добывают из семян подсолнечника, рапса, кукурузы, льна и многих других масличных культур.
Роль семян в природе и жизни человека
Биология семян и рассады
Выбор правильных семян
Прежде чем приступить к изучению способов выращивания семян и рассады, начните с правильных семян. Если вы намерены вести свою деятельность как сертифицированное органическое растение, вам необходимо использовать сертифицированные органические семена и саженцы, за некоторыми исключениями (см. «Требования к органическим веществам»).
Если вы намерены вести свою деятельность как сертифицированное органическое растение, вам необходимо использовать сертифицированные органические семена и саженцы, за некоторыми исключениями (см. «Требования к органическим веществам»).
Что необходимо семенам для прорастания?
Жизнеспособные семена — это живые существа. Для прорастания они должны содержать живую здоровую эмбриональную ткань. Все полностью развитые семена содержат зародыш и, у большинства видов растений, запас питательных веществ, завернутый в семенную оболочку. Семена обычно «пробуждаются» и прорастают, когда влажность почвы и температурные условия являются подходящими для их роста (Майлз и Браун, 2007). Каждый тип семян имеет свои индивидуальные потребности — найдите минутку и прочитайте об их конкретных требованиях к прорастанию.
Семенам нужна правильная среда для прорастания
Температура, влажность, воздух и освещение должны быть правильными для прорастания семян. Все семена имеют оптимальный температурный диапазон для прорастания (табл. 1). Минимальная температура – это самая низкая температура, при которой семена могут эффективно прорастать. Максимум – это самая высокая температура, при которой семена могут прорасти. Все, что выше или ниже этой температуры, может повредить семена или заставить их уйти в состояние покоя. При оптимальных температурах прорастание быстрое и равномерное.
1). Минимальная температура – это самая низкая температура, при которой семена могут эффективно прорастать. Максимум – это самая высокая температура, при которой семена могут прорасти. Все, что выше или ниже этой температуры, может повредить семена или заставить их уйти в состояние покоя. При оптимальных температурах прорастание быстрое и равномерное.
Все семена нуждаются в правильной влажности для запуска внутренних процессов, ведущих к прорастанию. В полевой почве это обычно составляет около 50-75 процентов полевой емкости. Для оптимального прорастания необходимы мелкозернистое семенное ложе и хороший контакт семян с почвой. Аэрация в почвенной среде обеспечивает хороший газообмен между прорастающим зародышем и почвой. Семена дышат так же, как и любой другой живой организм. Им нужен кислород, и они производят углекислый газ (CO 2 ). Этот углекислый газ должен иметь возможность удаляться от семян. Если почва или среда недостаточно аэрированы из-за переувлажнения или уплотнения, CO 2 не рассеется, и семена могут задохнуться.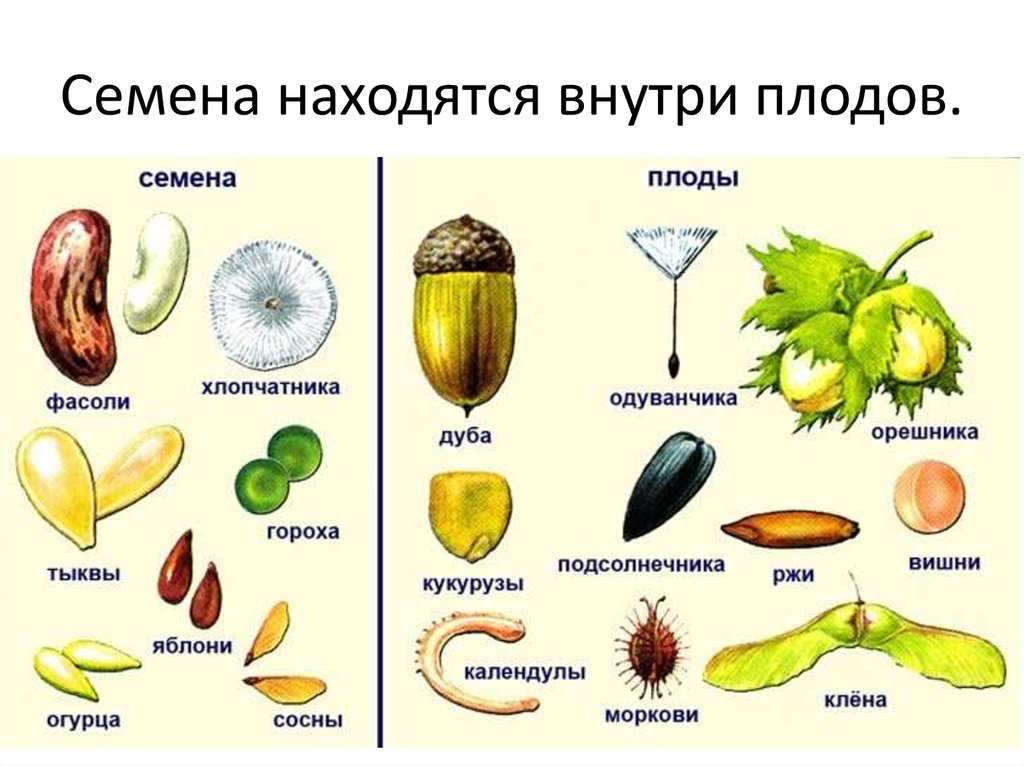
Не все семена одинаково нуждаются в свете. Большинство семян лучше всего прорастают в темных условиях и могут даже подавляться светом (например, Phacelia и Allium spp.). Однако некоторым видам (например, бегонии, примуле, колеусу) для прорастания необходим свет (Miles and Brown 2007). Не путайте требования к свету для семян с тем, что нужно рассаде. Все саженцы требуют солнечного света. Всходы станут длинноногими и хрупкими и не смогут раскрыть свой потенциал, если им не будет хватать света.
| Минимум (F) | Оптимальный диапазон (F) | Оптимальный (F) | Максимум (F) | |
|---|---|---|---|---|
| Свекла | 40 | 50-85 | 85 | 85 |
| Капуста | 40 | 45-95 | 85 | 100 |
| Цветная капуста | 40 | 45-85 | 80 | 100 |
| Сельдерей | 40 | 60-70 | 70 | 85 |
| Чард | 40 | 50-85 | 85 | 95 |
| Огурец | 60 | 60-95 | 95 | 105 |
| Баклажаны | 60 | 75-90 | 85 | 95 |
| Салат | 35 | 40-80 | 75 | 85 |
| Дыни | 60 | 75-95 | 90 | 100 |
| Лук | 35 | 50-95 | 75 | 95 |
| Петрушка | 40 | 50-85 | 75 | 90 |
| Перец | 60 | 65-95 | 85 | 95 |
| Тыква | 60 | 70-90 | 90 | 100 |
| Шпинат | 35 | 45-75 | 70 | 85 |
| Сквош | 60 | 70-95 | 95 | 100 |
| Помидор | 50 | 70-95 | 85 | 95 |
Температуру почвы следует измерять, вставив почвенный термометр на глубину 3-4 дюйма в поверхность почвы и отметив температуру.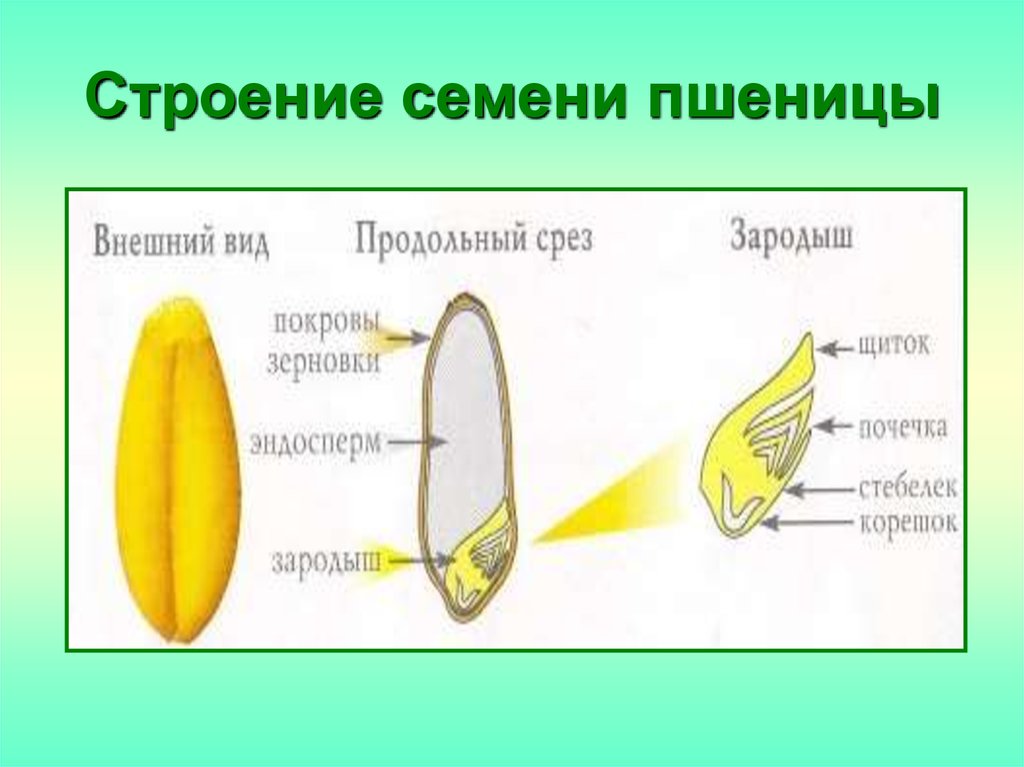 Адаптировано из Кембл и Масгроув (2006).
Адаптировано из Кембл и Масгроув (2006).
Покой семян
Некоторые жизнеспособные семена могут не прорасти. У многих семян развился период покоя (или сна). Покой семян – это состояние, препятствующее прорастанию даже в оптимальных условиях окружающей среды. Почему выгодно, чтобы не все семена прорастали при подходящих условиях? В природе ошеломляющая всхожесть защищает некоторые саженцы от возможных всплесков плохой погоды или травоядных, которые могут их съесть. Семена растений, которые лучше всего растут весной, самостоятельно прорастают только после того, как пройдут холодные зимние температуры.
Чтобы семена вышли из состояния покоя, мы должны разрушить их физические или химические факторы покоя. Семена могут иметь твердую или толстую семенную оболочку (физический покой). Это может быть нарушено путем замачивания или скарификации (царапания поверхности) семян. Другие семена имеют внутренние химические или метаболические условия, препятствующие прорастанию (химический покой). Факторы, влияющие на покой семян, включают присутствие некоторых растительных гормонов, в частности абсцизовой кислоты, подавляющей прорастание, и гиббереллина, прекращающего покой семян. Чтобы нарушить химический покой, вам, возможно, придется выщелачивать семена, использовать холодную/влажную стратификацию или огневую скарификацию. Например, мембрана семенной кожуры некоторых семян образует барьер, проницаемый для воды, но не для кислорода. Холодные температуры (50-59°F) позволяют кислороду проникать в семена, а теплые температуры препятствуют поглощению кислорода. Прохладные температуры также позволяют семенам переваривать часть своих пищевых запасов, давая им энергию. Помещение этих семян в холодильник на определенный период времени позволяет им получить достаточное количество кислорода и энергии для прорастания (Colorado Seed Laboratory 2009).
Факторы, влияющие на покой семян, включают присутствие некоторых растительных гормонов, в частности абсцизовой кислоты, подавляющей прорастание, и гиббереллина, прекращающего покой семян. Чтобы нарушить химический покой, вам, возможно, придется выщелачивать семена, использовать холодную/влажную стратификацию или огневую скарификацию. Например, мембрана семенной кожуры некоторых семян образует барьер, проницаемый для воды, но не для кислорода. Холодные температуры (50-59°F) позволяют кислороду проникать в семена, а теплые температуры препятствуют поглощению кислорода. Прохладные температуры также позволяют семенам переваривать часть своих пищевых запасов, давая им энергию. Помещение этих семян в холодильник на определенный период времени позволяет им получить достаточное количество кислорода и энергии для прорастания (Colorado Seed Laboratory 2009).
Этапы прорастания семян
- Набухание. Семя быстро впитывает воду, семенная кожура набухает и размягчается. Подумайте о семени гороха, которое вы замочили — внешняя оболочка семени становится мягкой и морщинистой от воды.
- Промежуточная или запаздывающая фаза. Во время этой фазы семя активирует свою внутреннюю физиологию, клетки дышат, и семя начинает вырабатывать белки и метаболизировать свои запасы пищи (MacKean n.d.).
- Появление корня и корня. Клетки начинают удлиняться и делиться, выводя корень и корешок из семени.
 Подумайте о семени гороха, которое вы замочили — внешняя оболочка семени становится мягкой и морщинистой от воды.
Подумайте о семени гороха, которое вы замочили — внешняя оболочка семени становится мягкой и морщинистой от воды.
Чтобы узнать, жизнеспособны ли ваши семена, проведите тест на всхожесть. Заверните семена во влажное бумажное полотенце, подождите 5-10 дней и посчитайте, сколько семян прорастет.
Иллюстрация 1: Этапы прорастания семян.
Если вы сохраняете семена прошлого года, подумайте вот о чем: срок службы семян можно сократить вдвое, увеличив влажность семян всего на 1 процент или повысив температуру хранения всего на несколько градусов. Простое эмпирическое правило заключается в том, что сумма температуры хранения (в градусах Фаренгейта) и относительной влажности в процентах не должна превышать 100. 0011
0011
Первичный корень, называемый корешком, появляется первым из семени. Первичный корень прикрепляет растение к земле и позволяет ему начать поглощать воду. После того, как корень впитает воду, из семени появится побег. У двудольных побег состоит из трех основных частей: семядолей (семенных листьев), участка побега под семядолями (гипокотиля) и участка побега над семядолями (эпикотиля). То, как росток появляется из почвы или среды для выращивания, следует двум основным закономерностям. У некоторых растений участок побега под семядолями удлиняется и образует крючок, протягивая семядоли и растущий кончик через почву. Как только он достигает поверхности, он выпрямляется и тянет семядоли и кончики побегов растущих проростков в воздух. Например, так прорастают бобы. Это называется эпигейным прорастанием. У других растений расширяется только участок над семядолями, оставляя семядоли под землей, где они вскоре разлагаются. Это называется гипогеозным прорастанием. Так, например, прорастает горох (Raven, Ray, and Eichhorn 2005).
Однодольные (односемянные листья)
У семян однодольных первичный корень защищен оболочкой (колеориза), которая сначала выталкивается из семени. Затем появляются листья проростков, покрытые защитной оболочкой, называемой колеоптилем (Raven, Ray, and Eichhorn, 2005).
Двудольные и однодольные растения
После появления всходов проросток растет медленно, в то время как запасающая ткань семени уменьшается. Вскоре у растения развивается разветвленная корневая система или стержневой корень. Затем появляются настоящие листья, похожие на листья взрослого растения. Эти листья, в отличие от семядолей, фотосинтезируют свет в энергию, позволяя растению расти и развиваться.
Управление оптимальным прорастанием и развитием проростков
Оптимизация прорастания
Мы знаем, что семенам для прорастания требуется оптимальное количество воды, кислорода, температуры и света. Если мы не создадим максимально оптимальную среду, растения будут прорастать медленно и неравномерно. Как правило, площадь теплицы ограничена, поэтому мы хотим, чтобы растения проросли как можно быстрее. Неравномерное прорастание также может вызвать проблемы. Если вам когда-либо приходилось пересаживать рассаду, половина из которой готова к посадке, а другая половина слишком мала, с корневыми комами, которые не могут легко выскользнуть из ячеек, вы поймете, почему.
Как правило, площадь теплицы ограничена, поэтому мы хотим, чтобы растения проросли как можно быстрее. Неравномерное прорастание также может вызвать проблемы. Если вам когда-либо приходилось пересаживать рассаду, половина из которой готова к посадке, а другая половина слишком мала, с корневыми комами, которые не могут легко выскользнуть из ячеек, вы поймете, почему.
Одним из распространенных способов достижения оптимальной температуры прорастания в средах для выращивания является использование матов для проращивания. Эти коврики позволяют устанавливать температуру в соответствии с требованиями семян. Например, перцы прорастают за 8 дней при 86°F, но для прорастания при 58°F требуется более 13 дней (Клуб хранителей семян Пенсильвании, н.д.).
Убедитесь, что вы поддерживаете оптимальную температуру для вашей культуры (см. Таблицу 1 выше). Также очень важно обеспечить циркуляцию воздуха, чтобы смягчить воздействие грибковых патогенов, вызывающих выпревание.
Развитие рассады
Оптимальная температура для выращивания рассады может отличаться от температуры для семян (таблица 2). Помните, что оптимальная температура будет стимулировать оптимальный рост. Вы можете контролировать температуру, чтобы контролировать высоту растений. Более низкие температуры обычно замедляют рост, а более теплые ускоряют рост.
Помните, что оптимальная температура будет стимулировать оптимальный рост. Вы можете контролировать температуру, чтобы контролировать высоту растений. Более низкие температуры обычно замедляют рост, а более теплые ускоряют рост.
| День (F) | Ночь (F) | Время (недели) | |
|---|---|---|---|
| Брокколи | 60-70 | 50-60 | 5-7 |
| Капуста | 60-70 | 50-60 | 5-7 |
| Цветная капуста | 60-70 | 50-60 | 5-7 |
| Сельдерей | 65-75 | 60-65 | 10-12 |
| Огурец | 70-75 | 60-65 | 3-4 |
| Баклажаны | 70-80 | 65-70 | 6-8 |
| Салат | 55-65 | 50-55 | 5-7 |
| Дыни | 70-80 | 65-70 | 3-4 |
| Лук | 60-65 | 55-60 | 10-12 |
| Перец | 65-75 | 60-65 | 6-8 |
| Сквош | 70-75 | 60-65 | 3-4 |
| Помидор | 65-75 | 60-65 | 5-7 |
Из Maynard and Hochmuth (2007).
Очень важно поддерживать хорошую циркуляцию воздуха и достаточную влажность. Как правило, полив должен быть более глубоким, чтобы обеспечить развитие корневой системы. Возможно, вам придется использовать разные шланги или шланги для полива семян и рассады, потому что каждый из них использует разное количество воды. Не забывайте внимательно следить и поливать растения по краям квартир. Они сохнут быстрее, чем средние. Однако чрезмерный полив может увеличить вероятность выпревания растений.
Созревание и закаливание семян
Этот последний шаг перед высадкой саженцев в поле постепенно подвергает их воздействию условий, в которых они будут находиться в поле. Этот процесс стимулирует растения накапливать запасы углеводов и питательных веществ и укреплять клеточные стенки, подвергая растения дневным и ночным колебаниям температуры, усиленному движению воздуха и ветру, уменьшению полива и полному освещению.
Закаливание рассады очень важно, особенно если ее высаживают в стрессовых условиях в начале сезона. Большинство рассады можно закалить, снизив температуру в теплице с помощью вентиляции. Уменьшенный полив также даст некоторый закаливающий эффект. Не допускайте чрезмерного увядания растений. Не закаляйте рассаду, уменьшая количество удобрений, так как это часто приводит к низкорослым растениям, которые плохо приживаются в поле. Некоторые цветоводы выносят растения на улицу за 5-7 дней до посадки. Это позволяет растению адаптироваться к внешним условиям еще в квартире. Растения, закаленные таким образом, часто имеют более высокие полевые показатели по сравнению с растениями, посаженными непосредственно из теплицы (Garton, Sikkema, Tomecek 19).97).
Большинство рассады можно закалить, снизив температуру в теплице с помощью вентиляции. Уменьшенный полив также даст некоторый закаливающий эффект. Не допускайте чрезмерного увядания растений. Не закаляйте рассаду, уменьшая количество удобрений, так как это часто приводит к низкорослым растениям, которые плохо приживаются в поле. Некоторые цветоводы выносят растения на улицу за 5-7 дней до посадки. Это позволяет растению адаптироваться к внешним условиям еще в квартире. Растения, закаленные таким образом, часто имеют более высокие полевые показатели по сравнению с растениями, посаженными непосредственно из теплицы (Garton, Sikkema, Tomecek 19).97).
Органические требования
Национальные органические стандарты требуют, чтобы производители использовали органически выращенные семена, однолетние саженцы и посадочный материал. Неорганически произведенные, необработанные семена и посадочный материал могут использоваться для производства органической культуры, когда эквивалентный органически произведенный сорт отсутствует в продаже.
Семена, обработанные запрещенными материалами, не допускаются. Каптан, тимет и подобные химические фунгициды не включены в национальный список и не разрешены к использованию. Пожалуйста, отнеситесь к этому серьезно. Если ваше семя покрыто розовым или оранжевым порошком, вероятно, оно запрещено. Возможно, мы не сможем сертифицировать ваш урожай, если вы используете семена, обработанные запрещенными материалами.
Семена, используемые для производства съедобных ростков, должны быть органическими — без исключений.
Коммерческая доступность
Первый шаг – определить, доступен ли эквивалентный органически произведенный сорт. По эквивалентному сорту ищите сопоставимые привычки роста, количество дней до созревания, устойчивость к насекомым и болезням, вкус и другие важные качества. Если подходящий органический эквивалентный сорт недоступен, задокументируйте, где вы пытались найти органические семена, так как это важно для ваших сертификационных записей. После того, как вы нашли источник конкретных эквивалентных органических семян, следующим шагом в определении коммерческой доступности является проверка того, соответствуют ли они форме, качеству и количеству.
- Форма: сортированная, гранулированная, обработанная горячей водой
- Качество: попробуйте небольшое количество в первый год, чтобы убедиться, что оно хорошо работает в ваших конкретных условиях; если единственные доступные органические семена имеют низкое качество, то покупка неорганических может быть приемлемой
- Количество: например, если вы хотите посадить 1 акр тыквы, а единственные доступные органические семена находятся в пакетах по 1 унции, тогда покупка неорганических может быть приемлемой
Документация и добросовестные усилия
Предварительное одобрение Пенсильванской сертифицированной органической организации для использования неорганических семян/посадочного материала не требуется. Соответствие проверяется в контексте плана органической системы, который проверяется во время ежегодной проверки. Образец неадекватной документации и отсутствие добросовестных усилий для получения органически выращенных семян и посадочного материала может рассматриваться как несоблюдение требований и может привести к тому, что Пенсильванская сертифицированная органическая продукция потребует предварительного одобрения в отношении вопросов коммерческой доступности в будущих циклах посадки. Документирование ваших добросовестных усилий по поиску подходящих органических семян/посадочного материала имеет решающее значение.
Документирование ваших добросовестных усилий по поиску подходящих органических семян/посадочного материала имеет решающее значение.
Источники органических семян
Ниже перечислены источники органических семян, предоставленные Pennsylvania Certified Organic (2011). Список также ведется Институтом обзора органических материалов (OMRI). Для получения более полного списка, включая кормовые, полевые и покровные культуры, семена и рассаду, посетите веб-сайт Pennsylvania Certified Organic.
Abundant Life Seeds
PO Box 157
Saginaw, OR 97472
Телефон: 541-767-9606
Сайт: www.abundantlifeseeds.com
Электронная почта: [email protected]
100% органические, все опыляются открытым способом; семена овощей, цветов и трав; чеснок и картофель
The Cook’s Garden
PO Box C5030
Warminster, PA 18974
Телефон: 800-457-9703
Сайт: www.cooksgarden.com
Электронная почта: cooksgarden@earthlink.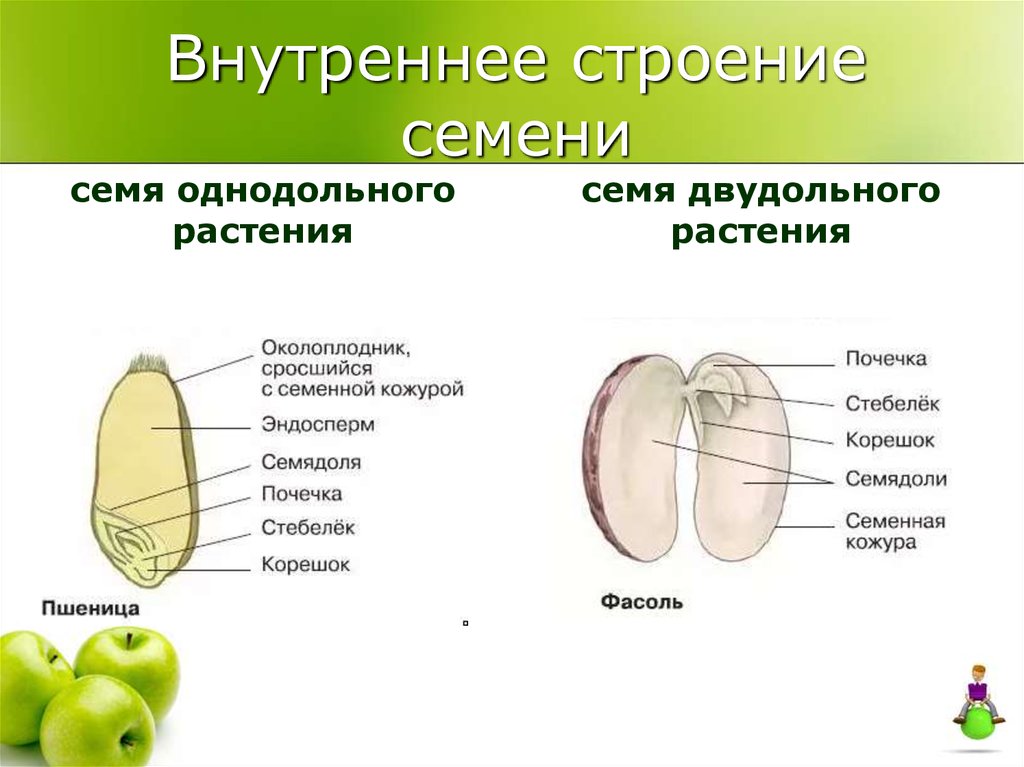 net 9026 , цветы и травы
net 9026 , цветы и травы
Экологические производители семян
PO Box 947
Albany, OR 97321-0354
Телефон: 541-928-5868
Веб-сайт: www.espseeds.com
Органические овощи, травы и цветы
Fedco Seeds
PO Box 520
Waterville, ME 04903
Телефон: 207-873-7333
Веб-сайт:
Органические овощи и цветы
Filaree Farm
182 Conconully Hwy
Okanogan, WA 98840
Телефон: 509-422-6940
Web: WWW.Filereefarm. и Ко
600 Mamaroneck Avenue
Harrison, NY 10528-1631
Телефон: 800-345-3787
Факс: 914-698-2857
Сайт: www.fredgloeckner.com
Органические семена овощей, трав и цветов Gardens
Alives !
500 Schenley Place
Lawrenceburg, IN 47025
Телефон: 513-354-1482
Веб-сайт: www.gardensalive.com
Органические семена для садовых растений и рассады, а также средства для борьбы с насекомыми и болезнями и средства по уходу за почвой
Harris Seeds 9022 Пол Роуд
PO Box 24966
Rochester, NY 14624-0966
Телефон: 800-544-7938
Веб: www. harisseeds.com
harisseeds.com
Некоторые органические овощи и травы
Высокие семена кожи.
Телефон: 802-472-6174
Веб-сайт: www.highmowingseeds.com
Высококачественные органические семена для более чем 500 сортов реликвии, свободно опыляемых и гибридных овощей, цветов, трав, картофеля, чеснока и покровных культур
Отборные семена Johnny’s
955 Benton Avenue
Winslow, ME 04901
Телефон: 877-JOHNNYS (877-564-6697)
Веб-сайт: www.johnnyseeds.com
Органические овощи, цветы и травы
The Maine Potato Lady 902 6 65
Guilford, ME 04443
Телефон: 207-343-2270
Сайт: www.mainepotatolady.com
Органический семенной картофель, лук-шалот, лук-севок, чеснок и покровные культуры, а также удобрения, почва и инокулянты для семян
Rohrer Семена
Почтовый ящик 250
Смоктаун, Пенсильвания 17576
Телефон: 717-299-2571
Веб-сайт: www.rohrerseeds.com
Органические семена овощей
Seeds of Change
3209 Richards Lane
Santa Fe, NM 87507
Веб-сайт: 888-739-726 seedofchange.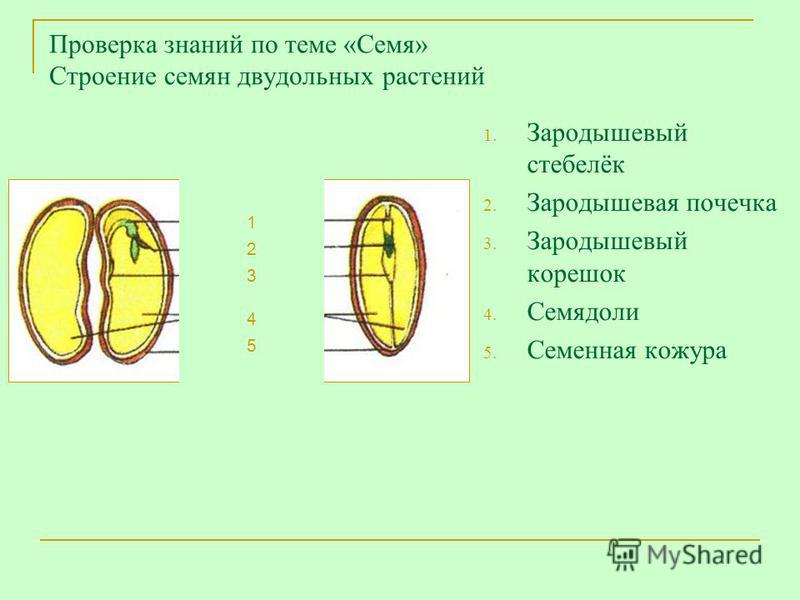 com
com
Органические цветы, травы, овощи, покровные культуры и растения клубники
Seed Savers Exchange
3094 North Winn Road
Decorah, IA 52101
Телефон: 563-382-5990
Интернет: www.seedsavers.org
Некоторые органические овощи, чеснок, травы, картофель и реликвии
Seedway
99 Industrial Road
Elizabethtown, PA 17022
Phone: 800-952-7333
Web: www.seedway.com
Some organic vegetables and herbs
Snow Seed Organic
21855 Rosehart Way
Salinas, CA 93908
Телефон: 831-758-9869
Веб-сайт: www.snowseedco.com
Многие органические овощи, в том числе салат-латук
Southern Exposure Seed Exchange
PO Box 460
Mineral, VA 23117
08-9 Телефон: 94-9480
Веб-сайт: www.southernexposure.com
Более 400 сортов сертифицированных органических реликвий и свободно опыляемых семян овощей, трав и цветов, а также луковицы чеснока и многолетнего лука
Территориальная семенная компания
Почтовый ящик 158
Cottage Grove, OR 97424
Телефон: 800-626-0866
Веб-сайт: www. territorialseed.com
territorialseed.com
Органические семена цветов, трав, овощей, чеснока и покровных культур, а также внесенные в список OMRI удобрения и почвоулучшители
Vitalis Органические семена
7 Harris Place
Salinas, CA 93901
Телефон: 831-262-7635
Веб-сайт: www.vitalisorganic.com
Органические семена овощей и трав, с акцентом на листья салата, шпинат, помидоры, перец, огурцы, кабачки и дыни
Wood Prairie Farm
49 Kinney Road
Bridgewater, ME 04735
Телефон: 800-829-9765
Сайт: www.woodprairie.com , RW, PH Sikkema и EJ Tomecek. Штепсельные рассады для переработки помидоров: производство, обработка и создание стенда. Онтарио, Канада: Министерство сельского хозяйства и сельских дел, 19 лет.97.
Кембл, Дж. и М. Масгроув. Температурные условия почвы для прорастания семян овощных культур. Alabama Cooperative Extension, 2006.
Маккин, Д.Г. «Биология растений: семена и прорастание», из «Ресурсов для преподавания биологии». 2010.
2010.
Мейнард Д. и Г. Хохмут. Справочник Нотта для овощеводов. Том. 5. Хобокен, Нью-Джерси: John Wiley and Sons, 2007.
Майлз, А. и М. Браун. Преподавание органического земледелия и садоводства: ресурсы для инструкторов. Санта-Крус: Ферма и сад Калифорнийского университета, 2007 г. 9.0005
Пенсильванский сертифицированный органический продукт. «Поставщики органических семян 2011 г.».
——. «Требования к семенам, посадочному материалу и саженцам в соответствии с правилами Министерства сельского хозяйства США».
Пенсильванский клуб хранителей семейных реликвий. «Прорастание семян и температура».
Рэйвен, П. Х., Ф. Э. Рэй и С. Э. Эйххорн. Биология растений. 7-е изд. Нью-Йорк: WH Freeman, 2005.
Образовательные программы по технологии семян. «Физиологический покой». Форт-Коллинз: Лаборатория семян Колорадо, 2009 г..
Подготовлено С. Тианной Дюпон, бывшим преподавателем устойчивого сельского хозяйства, Penn State Extension. Проверено Эльзой Санчес, Департамент садоводства штата Пенсильвания, и Деброй Брубейкер, сертифицированным органическим органом штата Пенсильвания.
Эта публикация была частично поддержана за счет средств Программы развития начинающих фермеров и владельцев ранчо Национального института продовольствия и сельского хозяйства Министерства сельского хозяйства США, грант № 2009-49400-05869.
Понимание биологии семян и рассады
Семена некоторых сельскохозяйственных культур, таких как помидоры, перец и брокколи, обычно сажают в тепличные лотки для выращивания рассады для последующей пересадки в поле.
Другие культуры, такие как сладкая кукуруза, фасоль и горох, чаще всего выращиваются путем посева семян прямо в поле. Понимание того, как семена прорастают и превращаются в здоровые саженцы, имеет решающее значение для успешного выращивания урожая. Это краткое изложение биологии семян и рассады, а также рекомендации по выращиванию здоровой рассады.
Выбор правильных семян
Немногие решения важнее для успешного выращивания овощей, чем выбор правильных семян. Семена должны быть высокого качества, чистыми и приобретаться у надежной семенной компании. Старайтесь выбирать сорта, которые хорошо адаптированы к местности и соответствуют предпочтениям местных потребителей. Сертифицированные органические производители обязаны использовать сертифицированные органические семена и саженцы, за некоторыми исключениями.
Старайтесь выбирать сорта, которые хорошо адаптированы к местности и соответствуют предпочтениям местных потребителей. Сертифицированные органические производители обязаны использовать сертифицированные органические семена и саженцы, за некоторыми исключениями.
Правильная среда для прорастания
Все полностью развитые семена содержат зародыш и, у большинства видов растений, запас питательных веществ, завернутый в семенную оболочку. Семена обычно «просыпаются» и прорастают, когда влажность почвы и температурные условия благоприятны для их роста. Но не все семена имеют одинаковые требования к прорастанию, поэтому важно знать, что нужно каждому типу семян.
Семенам для прорастания необходимы надлежащие условия температуры, влажности, воздуха и освещения. Все семена имеют оптимальный температурный диапазон для прорастания (табл. 1). Минимальная температура – это самая низкая температура, при которой семена могут эффективно прорастать. Максимум – это самая высокая температура, при которой семена могут прорасти. Все, что выше или ниже этой температуры, может повредить семена или заставить их уйти в состояние покоя. При оптимальных температурах прорастание быстрое и равномерное.
Все, что выше или ниже этой температуры, может повредить семена или заставить их уйти в состояние покоя. При оптимальных температурах прорастание быстрое и равномерное.
Для прорастания всем семенам нужна влага и воздух. Очень важен хороший контакт семян с почвой. Для прямого посева в поле рекомендуется мелкозернистое семенное ложе с небольшим уплотнением. Для тепличных пересадок беспочвенная среда должна быть влажной и рыхлой. Почва или среда для выращивания должны содержать достаточно влаги, чтобы семена могли впитывать воду, чтобы начать процесс прорастания. Но если почва или среда слишком влажные или слишком уплотненные, вокруг семени не будет достаточно воздуха, чтобы «дышать». Как и любому другому живому организму, семенам для дыхания необходим кислород, и этот кислород поступает из воздуха в почве.
Не все семена одинаково нуждаются в свете. Большинство семян лучше всего прорастают в темных условиях и даже могут быть подавлены светом. Некоторым другим семенам для прорастания нужен свет. Но как только семена проросли и проросли через поверхность почвы или питательной среды в виде проростков, всем им для роста необходим солнечный свет. В таблице 1 приведены оптимальные температуры почвы для прорастания обычных овощных культур.
Но как только семена проросли и проросли через поверхность почвы или питательной среды в виде проростков, всем им для роста необходим солнечный свет. В таблице 1 приведены оптимальные температуры почвы для прорастания обычных овощных культур.
| Овощи | Минимум (°F) | Оптимальный диапазон (°F) | Оптимум (°F) | Максимум (°F) |
|---|---|---|---|---|
| 40 | 50-85 | 85 | 85 | |
| Фасоль | 60 | 60-85 | 80 | 95 |
| Капуста | 40 | 45-95 | 85 | 100 |
| Цветная капуста | 40 | 45-85 | 80 | 100 |
| Сельдерей | 40 | 60-70 | 70 | 85 |
| Чард | 40 | 50-85 | 85 | 95 |
| Огурец | 60 | 60-95 | 95 | 105 |
| Баклажаны | 60 | 75-90 | 85 | 95 |
| Салат | 35 | 40-80 | 75 | 85 |
| Дыни | 60 | 75-95 | 90 | 100 |
| Лук | 35 | 50-95 | 75 | 95 |
| Петрушка | 40 | 50-85 | 75 | 90 |
| Горох | 40 | 40-75 | 75 | 85 |
| Перец | 60 | 65-95 | 85 | 95 |
| Тыква | 60 | 70-90 | 90 | 100 |
| Шпинат | 35 | 45-75 | 70 | 85 |
| Сквош | 60 | 70-95 | 95 | 100 |
| Сладкая кукуруза | 50 | 60-95 | 95 | 105 |
| Помидор | 50 | 70-95 | 85 | 95 |
Шаги от семени к проростку
Когда сухое семя вступает в контакт с влажной почвой или средой для выращивания, семя начинает поглощать воду через семенную оболочку. По мере того, как он поглощает больше воды, семя расширяется, и семенная кожура трескается. Зародыш внутри семени состоит из маленького побега и маленького корня. Корень появляется первым из семени. По мере роста он прикрепляет растение к земле и начинает поглощать воду через корень. После того, как корень впитает воду, из семени начинает прорастать побег.
По мере того, как он поглощает больше воды, семя расширяется, и семенная кожура трескается. Зародыш внутри семени состоит из маленького побега и маленького корня. Корень появляется первым из семени. По мере роста он прикрепляет растение к земле и начинает поглощать воду через корень. После того, как корень впитает воду, из семени начинает прорастать побег.
Большинство овощей, которые мы выращиваем, являются двудольными. Двудольные имеют два семенных листа в побеге, которые появляются из прорастающего семени. Помидоры, перец, капуста, свекла, салат, фасоль, огурцы и кабачки — все это двудольные. Некоторые овощи относятся к однодольным, в том числе сладкая кукуруза, лук и спаржа. Однодольные имеют один семенной лист в формирующемся побеге. Как только побег с одним или двумя семенными листочками появляется из почвы или среды для выращивания, мы называем растение проростком. Все еще довольно хрупкий на этой стадии, молодой саженец уязвим для болезней и стрессов окружающей среды, таких как высокие температуры и сухая почва. Обеспечение оптимальных условий для роста гарантирует быстрый и равномерный рост молодых саженцев.
Обеспечение оптимальных условий для роста гарантирует быстрый и равномерный рост молодых саженцев.
Управление для оптимального прорастания и развития рассады
Проверка сохраненных семян на всхожесть
Семена, купленные в течение года, когда они должны быть посажены, редко не прорастают. Однако семена часто хранятся из года в год, и если они хранятся неправильно или слишком много лет, они могут потерять силу и плохо прорасти при посадке. Простой тест на всхожесть может показать, жизнеспособны ли сохраненные семена. Для проверки семян на всхожесть отсчитывают не менее двадцати пяти семян. Слегка заверните семена во влажное бумажное полотенце, держите бумажное полотенце влажным, но не мокрым в течение пяти-десяти дней. Разверните бумажное полотенце и посчитайте, сколько семян проросло. Если меньше 85-9Проросло 0% семян, остальные лучше выбросить и приобрести новые семена.
Равномерная всхожесть
Мы знаем, что семенам нужны надлежащие условия для быстрого прорастания.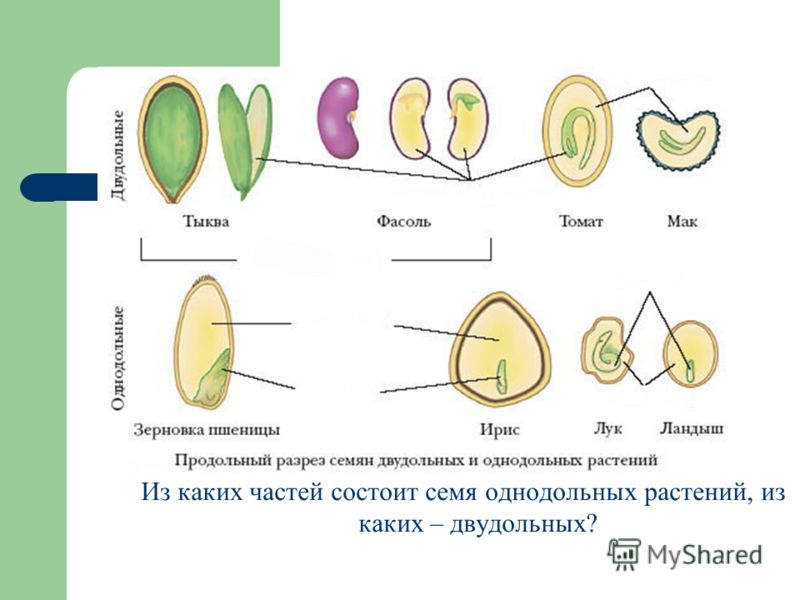 Независимо от того, сажают ли семена в лотки в теплице или прямо в поле, цель состоит в том, чтобы все семена прорастали примерно в одно и то же время и росли с одинаковой скоростью. Равномерным лотком с рассадой или однородным полем с рассадой легче управлять, и это приведет к лучшему урожаю. Неравномерное прорастание из-за медленного роста, различий во влажности или температуре почвы или глубине заделки семян может привести к получению всходов разного размера. Особенно это может вызвать проблемы при пересадке лотка с рассадой; половина готова к посадке в поле, а другая половина слишком мала, с корневыми комами, которые не могут легко выскользнуть из ячеек лотка.
Независимо от того, сажают ли семена в лотки в теплице или прямо в поле, цель состоит в том, чтобы все семена прорастали примерно в одно и то же время и росли с одинаковой скоростью. Равномерным лотком с рассадой или однородным полем с рассадой легче управлять, и это приведет к лучшему урожаю. Неравномерное прорастание из-за медленного роста, различий во влажности или температуре почвы или глубине заделки семян может привести к получению всходов разного размера. Особенно это может вызвать проблемы при пересадке лотка с рассадой; половина готова к посадке в поле, а другая половина слишком мала, с корневыми комами, которые не могут легко выскользнуть из ячеек лотка.
В теплице одним из способов добиться быстрого и равномерного прорастания является использование ковриков для проращивания под лотками. Эти коврики позволяют устанавливать температуру в соответствии с требованиями семян. Например, перцы прорастают за 8 дней при 86°F, но для прорастания при 58°F требуется более 13 дней. Убедитесь, что вы поддерживаете оптимальную температуру для вашей культуры (см. Таблицу 1). Обеспечение хорошей циркуляции воздуха во время прорастания и раннего выращивания рассады поможет контролировать болезни на этой ранней стадии.
Убедитесь, что вы поддерживаете оптимальную температуру для вашей культуры (см. Таблицу 1). Обеспечение хорошей циркуляции воздуха во время прорастания и раннего выращивания рассады поможет контролировать болезни на этой ранней стадии.
Мы не можем контролировать условия в поле, как в теплице, но мы все же можем принять меры, чтобы семена, посаженные прямо в поле, прорастали равномерно. Мелкозернистое семенное ложе обеспечивает хорошие условия для роста, достаточный контакт семян с почвой и возможность посадки на одинаковую глубину. Посадка при температуре почвы, близкой к оптимальной, ускорит прорастание и появление всходов. Иногда в спешке весенней посадки семена высевают в слишком холодную почву. Это может привести к медленному прорастанию, ослаблению и заболеванию всходов и даже к гибели растений. Посадку лучше отложить до прогрева почвы.
Развитие рассады
Оптимальная температура для выращивания рассады может отличаться от оптимальной температуры прорастания.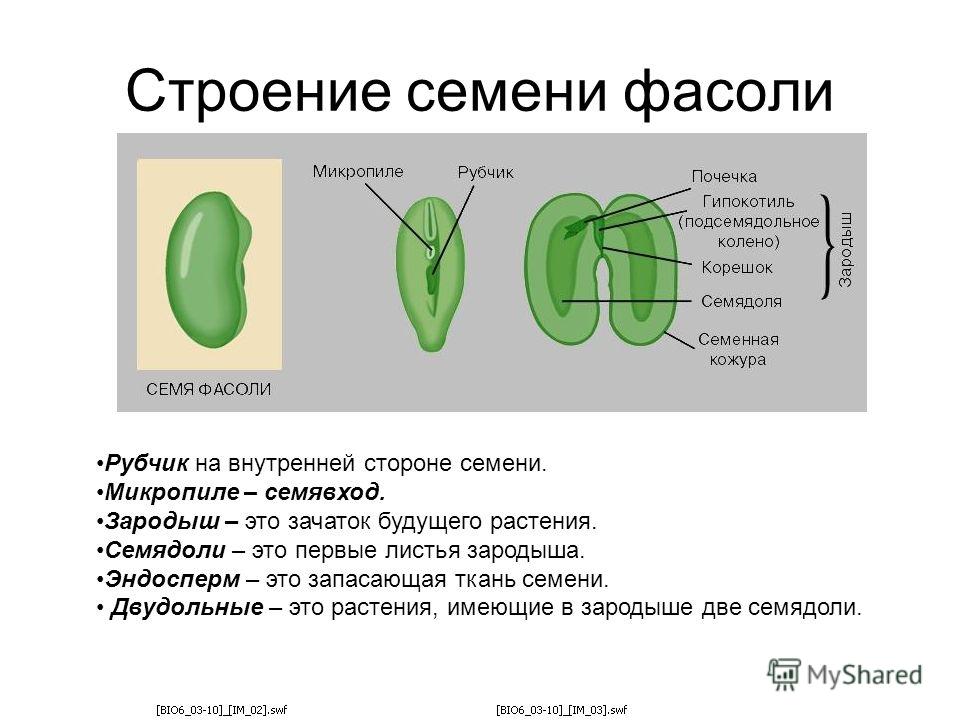 В таблице 2 показан диапазон дневных и ночных температур, оптимальных для выращивания рассады в теплице, где можно контролировать температуру. Более низкие температуры обычно замедляют рост, а более теплые ускоряют рост рассады.
В таблице 2 показан диапазон дневных и ночных температур, оптимальных для выращивания рассады в теплице, где можно контролировать температуру. Более низкие температуры обычно замедляют рост, а более теплые ускоряют рост рассады.
Все саженцы нуждаются в достаточном количестве света для роста. Если уровень освещенности низкий или сеянцам слишком тесно по мере роста, стебли будут вытягиваться, поскольку растения ищут больше света, что приводит к слабым, «длинноногим» пересадкам. Подумайте о дополнительном освещении, если уровень освещенности в теплице низкий.
| Овощи | День ( o F) | Ночь ( o F) | Время (недели) | 60-70 | 50-60 | 5-7 |
|---|---|---|---|
| Капуста | 60-70 | 50-60 | 5-7 |
| Цветная капуста | 60-70 | 50-60 | 5-7 |
| Сельдерей | 65-75 | 60-65 | 10-12 |
| Огурец | 70-75 | 60-65 | 3-4 |
| Баклажаны | 70-80 | 65-70 | 6-8 |
| Салат | 55-65 | 50-55 | 5-7 |
| Дыни | 70-80 | 65-70 | 3-4 |
| Лук | 60-65 | 55-60 | 10-12 |
| Перец | 65-75 | 60-65 | 6-8 |
| Сквош | 70-75 | 60-65 | 3-4 |
| Помидор | 65-75 | 60-65 | 5-7 |
Созревание и закаливание рассады
Продолжительность времени, в течение которого рассада должна расти в теплице, прежде чем она станет достаточно большой для пересадки в поле, зависит от культуры.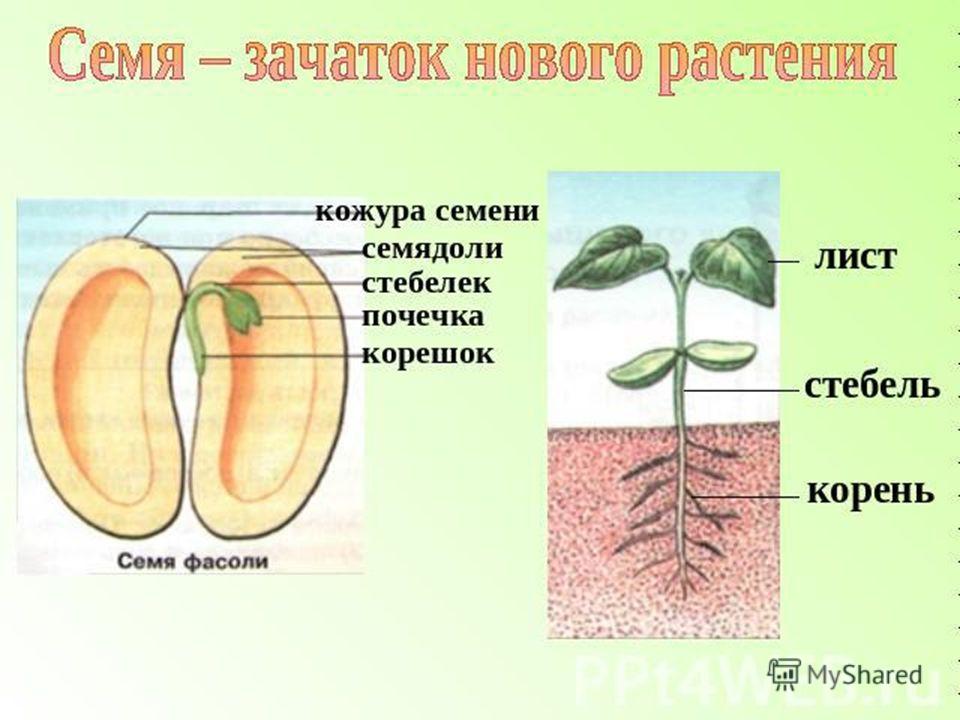 Для выращивания рассады помидоров и перца может потребоваться от пяти до семи недель, в то время как огурцы и кабачки готовы к пересадке через три-четыре недели в теплице. Но все трансплантаты должны быть закалены перед тем, как отправиться из теплицы в поле, иначе они будут повреждены более суровыми условиями. Чтобы закалить сеянцы, постепенно подвергайте их воздействию условий, которые у них будут в поле. Большинство рассады можно закалить, снизив температуру в теплице и/или сократив полив. Растения могут проявлять некоторые признаки увядания, но не допускайте их чрезмерного увядания.
Для выращивания рассады помидоров и перца может потребоваться от пяти до семи недель, в то время как огурцы и кабачки готовы к пересадке через три-четыре недели в теплице. Но все трансплантаты должны быть закалены перед тем, как отправиться из теплицы в поле, иначе они будут повреждены более суровыми условиями. Чтобы закалить сеянцы, постепенно подвергайте их воздействию условий, которые у них будут в поле. Большинство рассады можно закалить, снизив температуру в теплице и/или сократив полив. Растения могут проявлять некоторые признаки увядания, но не допускайте их чрезмерного увядания.
Через день или два, если позволяет погода, поставьте лотки вне теплицы на пять-семь дней до посадки. Если очень жарко и солнечно, обеспечьте рассаде тень на первые день-два. Растения, закаленные таким образом, лучше переносят пересадку и продолжают расти в поле без перерыва.
Для получения дополнительной информации см. «Биология семян и рассады».
При финансовой поддержке проекта Министерства сельского хозяйства США по гранту на специальные культуры ME № 44166076 – «Инновации в области устойчивого производства и борьбы с вредителями для молодых и испаноязычных/латиноамериканских производителей специальных культур следующего поколения»
прорастание | Описание, процесс, схема, этапы, типы и факты
семядоли и прорастание
См. все среды
все среды
- Связанные темы:
- ростковая трубка криптогейное прорастание эпигейное прорастание гипогеозное прорастание апрессорий
Просмотреть весь связанный контент →
Резюме
Прочтите краткий обзор этой темы
прорастание , прорастание семени, споры или другого репродуктивного тела, обычно после периода покоя. Поглощение воды, течение времени, охлаждение, согревание, наличие кислорода и воздействие света могут влиять на инициирование процесса.
В процессе прорастания семян вода поглощается зародышем, что приводит к регидратации и расширению клеток. Вскоре после начала поглощения воды, или впитывания, увеличивается частота дыхания и возобновляются различные обменные процессы, приостановившиеся или значительно сниженные в период покоя. Эти явления связаны со структурными изменениями в органеллах (перепончатых телах, связанных с обменом веществ), в клетках зародыша.
Дополнительные сведения по этой теме
семена: прорастание
Покой имеет как минимум три функции: (1) немедленное прорастание должно быть предотвращено даже при оптимальных обстоятельствах. ..
..
Прорастание иногда происходит в начале процесса развития; зародыш мангрового дерева ( Rhizophora ) развивается внутри семязачатка, выталкивая набухший рудиментарный корень через еще прикрепленный цветок. У гороха и кукурузы (кукурузы) семядоли (семенные листья) остаются под землей (например, гипогеальное прорастание), в то время как у других видов (фасоли, подсолнечника и т. д.) гипокотиль (эмбриональный стебель) растет на несколько дюймов над землей, неся семядоли. на свет, при котором они становятся зелеными и часто листовидными (например, эпигеальное прорастание).
Период покоя у некоторых семян кратковременный, например, у некоторых недолговечных однолетних растений. После рассеивания и при соответствующих условиях окружающей среды, таких как подходящая температура и доступ к воде и кислороду, семя прорастает, и зародыш возобновляет рост.
Семена многих видов прорастают не сразу после воздействия в целом благоприятных для роста растений условий, а требуют «нарушения» покоя, что может быть связано с изменением семенных оболочек или с состоянием самого зародыша. Обычно у зародыша нет врожденного покоя, и он развивается после того, как семенная оболочка удалена или достаточно повреждена, чтобы внутрь могла проникнуть вода. Прорастание в таких случаях зависит от гниения или истирания семенной кожуры в кишечнике животного или в почве. Ингибиторы прорастания должны быть либо вымыты водой, либо ткани, содержащие их, должны быть разрушены, прежде чем прорастание может произойти. Механическое ограничение роста зародыша характерно только для видов с толстой и прочной семенной оболочкой. Затем прорастание зависит от ослабления оболочки в результате истирания или разложения.
Обычно у зародыша нет врожденного покоя, и он развивается после того, как семенная оболочка удалена или достаточно повреждена, чтобы внутрь могла проникнуть вода. Прорастание в таких случаях зависит от гниения или истирания семенной кожуры в кишечнике животного или в почве. Ингибиторы прорастания должны быть либо вымыты водой, либо ткани, содержащие их, должны быть разрушены, прежде чем прорастание может произойти. Механическое ограничение роста зародыша характерно только для видов с толстой и прочной семенной оболочкой. Затем прорастание зависит от ослабления оболочки в результате истирания или разложения.
У многих семян зародыш не может прорасти даже в подходящих условиях, пока не пройдет определенный период времени. Это время может потребоваться для продолжения зародышевого развития в семени или для некоторого необходимого завершающего процесса, известного как дозревание, природа которого остается неясной.
Оформите подписку Britannica Premium и получите доступ к эксклюзивному контенту.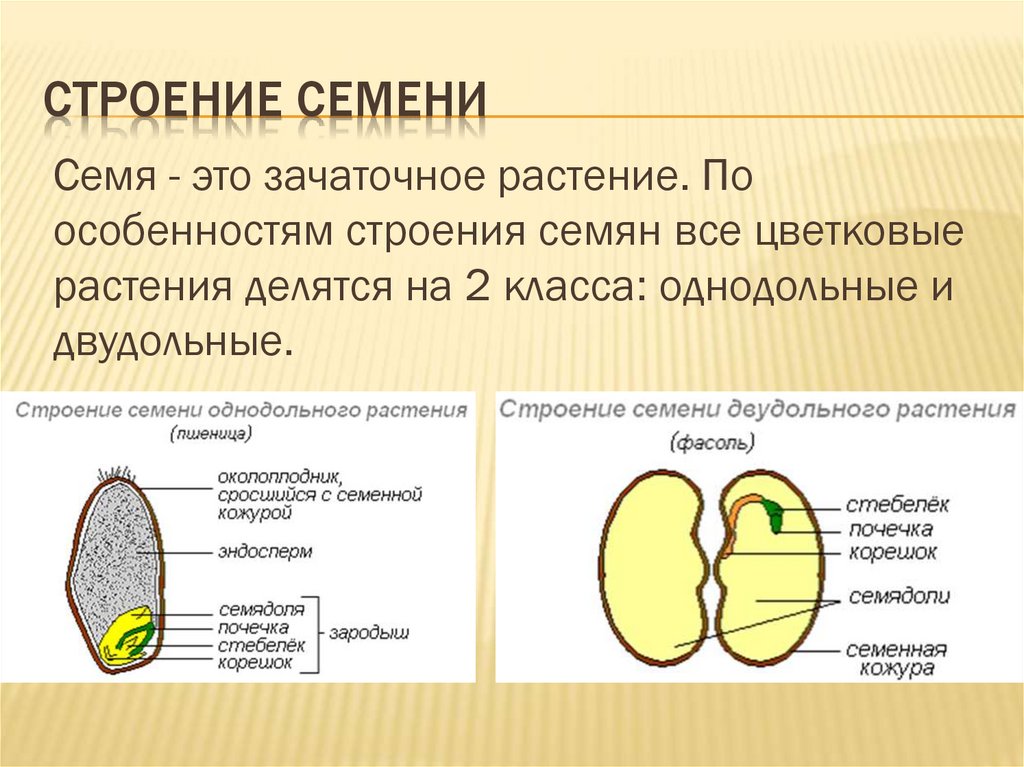 Подпишитесь сейчас
Подпишитесь сейчас
Семена многих растений, которые переносят холодные зимы, не прорастут, если они не пройдут период низкой температуры, обычно несколько выше нуля. В противном случае всходы не прорастают или сильно задерживаются, а ранний рост сеянцев часто бывает ненормальным. (Эта реакция семян на охлаждение имеет аналогию в контроле температуры покоя почек.) У некоторых видов прорастанию способствует воздействие света соответствующей длины волны. В других случаях свет подавляет прорастание. Для семян некоторых растений прорастанию способствует красный свет и тормозится свет с большей длиной волны в «дальнем красном» диапазоне спектра. Точное значение этой реакции пока неизвестно, но это может быть средством приведения времени прорастания в соответствие с сезоном года или определения глубины залегания семян в почве. Чувствительность к свету и требования к температуре часто взаимодействуют друг с другом, причем при определенных температурах потребность в свете полностью теряется.
Активный рост зародыша, за исключением набухания в результате набухания, обычно начинается с появления первичного корня, известного как корешок, из семени, хотя у некоторых видов (например, у кокоса) побег, или перышко, появляется первым. Ранний рост зависит в основном от экспансии клеток, но уже через короткое время начинается деление клеток в корешке и молодом побеге, а затем рост и дальнейшее формирование органов (органогенез) основаны на обычном сочетании увеличения числа клеток и укрупнения отдельных клеток. .
Пока росток не станет самостоятельным в плане питания, он зависит от запасов, обеспечиваемых родительским спорофитом. У покрытосеменных эти запасы находятся в эндосперме, в остаточных тканях семязачатка или в теле зародыша, обычно в семядолях. У голосеменных пищевые вещества содержатся преимущественно в женском гаметофите. Поскольку запасные материалы частично находятся в нерастворимой форме — в виде зерен крахмала, белковых гранул, капель липидов и т. п. — большая часть раннего метаболизма проростка связана с мобилизацией этих материалов и доставкой или перемещением продуктов в активные области.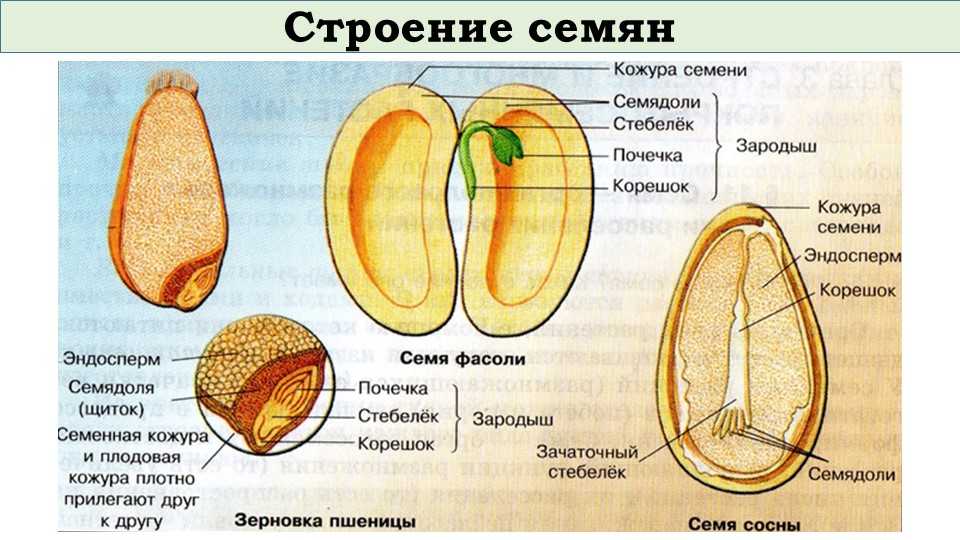 Запасы вне зародыша перевариваются ферментами, выделяемыми зародышем, а в некоторых случаях и специальными клетками эндосперма.
Запасы вне зародыша перевариваются ферментами, выделяемыми зародышем, а в некоторых случаях и специальными клетками эндосперма.
У некоторых семян (например, у клещевины) поглощение питательных веществ из запасов происходит через семядоли, которые позже расширяются на свету и становятся первыми органами, активными при фотосинтезе. Когда запасы накапливаются в самих семядолях, эти органы могут уменьшаться после прорастания и отмирать или вырабатывать хлорофилл и становиться фотосинтезирующими.
Факторы окружающей среды играют важную роль не только в определении ориентации проростка во время его укоренения, но также и в контроле некоторых аспектов его развития. Реакция сеянца на гравитацию очень важна. Корень, который обычно врастает в почву, называется положительно геотропным. Молодой побег, или перышко, считается отрицательно геотропным, поскольку он удаляется от почвы; он поднимается за счет удлинения либо гипокотиля, области между корешком и семядолями, либо эпикотиля, сегмента выше уровня семядолей.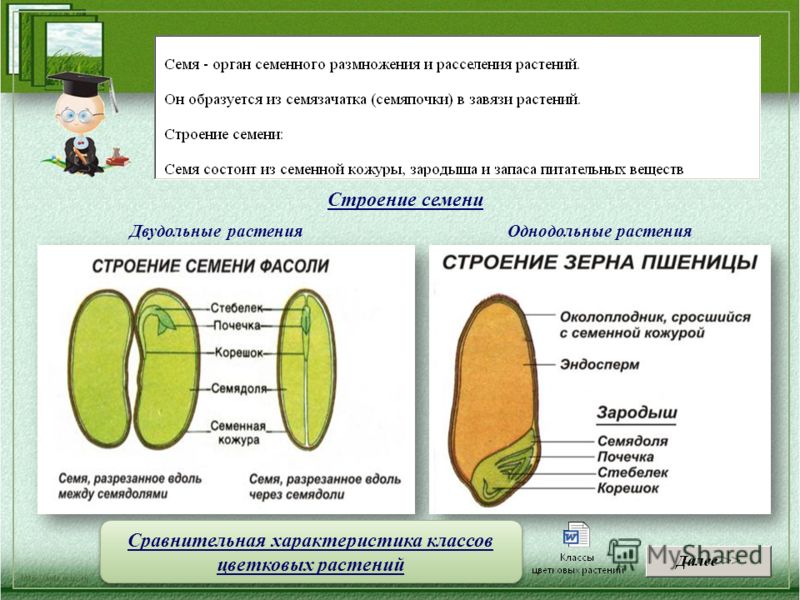 Если гипокотиль удлинен, семядоли выносятся из почвы. Если эпикотиль удлиняется, семядоли остаются в почве.
Если гипокотиль удлинен, семядоли выносятся из почвы. Если эпикотиль удлиняется, семядоли остаются в почве.
Свет влияет как на ориентацию ростка, так и на его форму. Когда семя прорастает под поверхностью почвы, перышко может появиться согнутым, защищая таким образом его нежную верхушку, только для того, чтобы выпрямиться на свету (искривление сохраняется, если побег выходит в темноту). Соответственно молодые листья перышка у таких растений, как фасоль, не расширяются и не становятся зелеными, кроме как после воздействия света. Известно, что эти адаптивные реакции регулируются реакциями, в которых участвует светочувствительный пигмент фитохром. У большинства сеянцев побеги проявляют сильное притяжение к свету или положительный фототропизм, который наиболее заметен, когда источник света направлен с одного направления. В сочетании с реакцией на гравитацию этот положительный фототропизм увеличивает вероятность того, что надземные части растения попадут в среду, наиболее благоприятную для фотосинтеза.
Q2 Представьте, что все семена, произведенные растением, попадают под одно и то же растение и прорастают в…
Перейти к
- Вопросы с множественным выбором (MCQ)
- Вопросы с краткими ответами (SA)
- Вопросы с длинными ответами (LA)
- Листок
- Цветок
- Клетка — структура и функции
- Пищеварительная система
- Дыхательная система
- Кровеносная система
- Здоровье и гигиена
- Среда обитания и адаптация
Главная > Селина Солюшнс Класс 6 Биология > Глава 2 — Цветок > Упражнение: Вопросы с длинными ответами (LA) > Вопрос 2
Вопрос 2 Вопросы с подробным ответом (LA)
Q2) Представьте, что все семена, произведенные растением, попадают под одно и то же растение и прорастают в новые растения. Укажите любые две проблемы, с которыми столкнутся новые заводы.
Укажите любые две проблемы, с которыми столкнутся новые заводы.
Ответ:
Решение:
Если все семена растения попадут под одно и то же растение и прорастут в новые растения, то в этой ситуации растения столкнутся со следующими проблемами:
- Большое количество растений будет расти в одном и том же месте, которое будет давать растениям лишь ограниченное количество воды и минералов.
- Они не получают достаточного количества воздуха и солнечного света, необходимого для нормального роста растения.
Стенограмма видео
«Здравствуйте, студенты, добро пожаловать на занятие по решению домашних заданий на лидо. Мы рассмотрим этот вопрос. Это к проблемам, с которыми столкнется новый план. Итак, учащиеся в этом вопросе. Нам нужно упомянуть две проблемы, с которыми столкнется новое растение, это оно должно быть в нем. Это растение или любое дерево и согласно вопрос представьте себе, что все семена, произведенные растением, попадают под один и тот же план.Так что вы можете видеть эту желтую часть, это семена, которые попали под один и тот же фронт Хорошо.Так вот как эти семена попадают под один и тот же план растение. Так что же будет, когда попадут под тот же план? Они прорастут новой кровью. Таков сценарий. новые растения проблема лемс, который будет за счет новых заводов, в первую очередь будет нехватка места.
Как видно из приведенной здесь схемы, здесь не хватает места, потому что здесь расстояние между двумя маленькими растениями или растениями очень меньше. Итак, места не хватает, а зачем нам пространство? Место нужно для роста листьев.
Повторяю, пространство необходимо для роста корней.
Точно так же, как есть последнее количество растений, растущих с той же скоростью. Они также будут ограничены в воде.
и минералы
Растению, потому что из-за нехватки места меньше места для роста корня. И именно поэтому будет обеспечено ограниченное количество воды и минералов, или растение будет получать ограниченное количество воды и минеральных веществ во вторую очередь.
Это растение или любое дерево и согласно вопрос представьте себе, что все семена, произведенные растением, попадают под один и тот же план.Так что вы можете видеть эту желтую часть, это семена, которые попали под один и тот же фронт Хорошо.Так вот как эти семена попадают под один и тот же план растение. Так что же будет, когда попадут под тот же план? Они прорастут новой кровью. Таков сценарий. новые растения проблема лемс, который будет за счет новых заводов, в первую очередь будет нехватка места.
Как видно из приведенной здесь схемы, здесь не хватает места, потому что здесь расстояние между двумя маленькими растениями или растениями очень меньше. Итак, места не хватает, а зачем нам пространство? Место нужно для роста листьев.
Повторяю, пространство необходимо для роста корней.
Точно так же, как есть последнее количество растений, растущих с той же скоростью. Они также будут ограничены в воде.
и минералы
Растению, потому что из-за нехватки места меньше места для роста корня. И именно поэтому будет обеспечено ограниченное количество воды и минералов, или растение будет получать ограниченное количество воды и минеральных веществ во вторую очередь.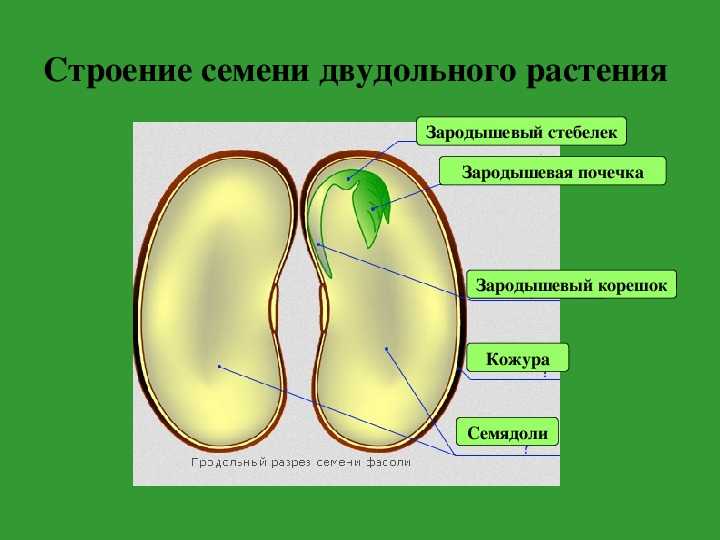 Они также не получат, они не получат его. Достаточное количество – это достаточное количество воздуха и солнечного света, а для чего им необходим солнечный свет, необходимый для процесса фотосинтеза через него и выщелачивания собственной пищи. Итак, как вы можете видеть, сосредоточьтесь на этом растении, вот основные планы, о которых я слышал от позднего, чтобы получить достаточное количество солнечного света. Нет, потому что Ял уже под улицей. Вот почему, поскольку они будут получать достаточное количество солнечного света. Они не вырастут до нужного уровня или не будет должной группы. Так что нормального роста нет. Так это этот провод? Это две проблемы, с которыми столкнется новое растение при нехватке места для бульона: они будут конкурировать, новое растение не получит все питательные вещества, а во-вторых, они не получают достаточного количества воздуха и солнечного света. так что нет нормального роста. Вот как мы пытаемся ответить на этот вопрос. Надеюсь, вы поняли решение. Если у вас есть какие-либо вопросы, оставьте комментарий ниже и подпишитесь на этот канал на регулярной основе.
Они также не получат, они не получат его. Достаточное количество – это достаточное количество воздуха и солнечного света, а для чего им необходим солнечный свет, необходимый для процесса фотосинтеза через него и выщелачивания собственной пищи. Итак, как вы можете видеть, сосредоточьтесь на этом растении, вот основные планы, о которых я слышал от позднего, чтобы получить достаточное количество солнечного света. Нет, потому что Ял уже под улицей. Вот почему, поскольку они будут получать достаточное количество солнечного света. Они не вырастут до нужного уровня или не будет должной группы. Так что нормального роста нет. Так это этот провод? Это две проблемы, с которыми столкнется новое растение при нехватке места для бульона: они будут конкурировать, новое растение не получит все питательные вещества, а во-вторых, они не получают достаточного количества воздуха и солнечного света. так что нет нормального роста. Вот как мы пытаемся ответить на этот вопрос. Надеюсь, вы поняли решение. Если у вас есть какие-либо вопросы, оставьте комментарий ниже и подпишитесь на этот канал на регулярной основе.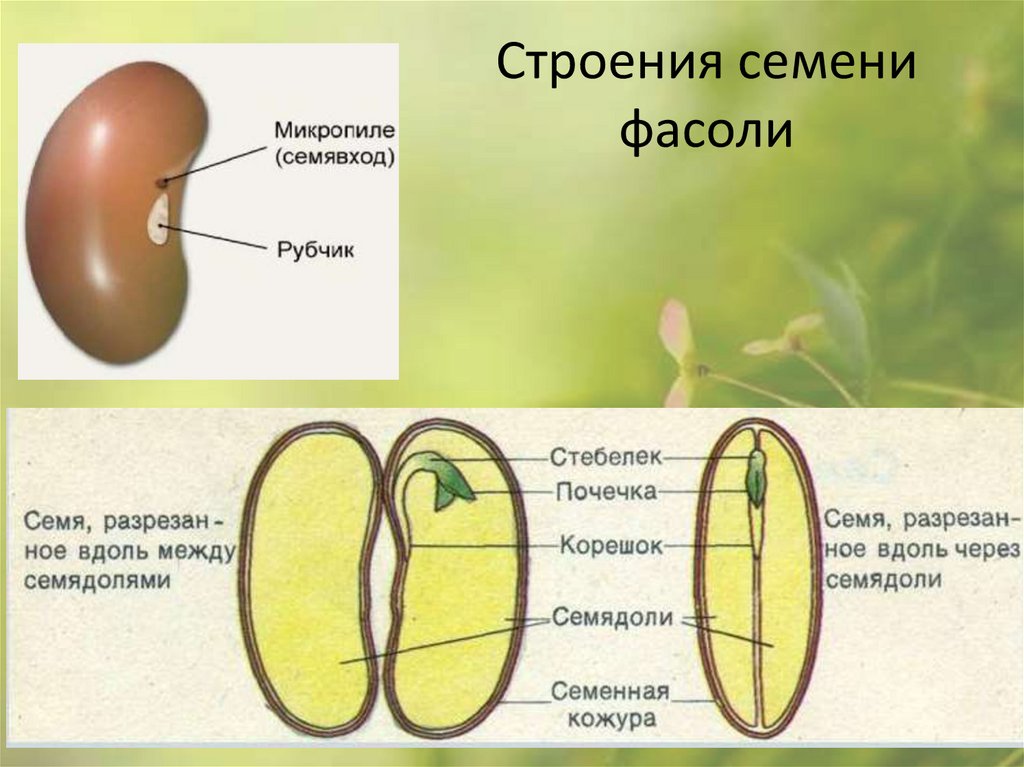 Хорошо. Спасибо.
Хорошо. Спасибо.
Связанные вопросы
Q1) Что подразумевается под опылением? Назовите два типа опыления.
Q3) Что такое цветок? Нарисуйте типичный цветок и подпишите его разные части.
Q4) С помощью подходящей размеченной схемы опишите строение семени двудольного растения.
Q5) Дайте определение всхожести? Назовите два типа прорастания. Объясните на примерах
Q6) Какие три условия необходимы для прорастания семян? Как бы вы продемонстрировали…
Q7) Назовите основное различие между гипогеальным и эпигеальным прорастанием.
Фейсбук WhatsApp
Копировать ссылку
Было ли это полезно?
Упражнения
Вопросы с несколькими вариантами ответов (MCQ)
Вопросы с короткими ответами (SA)
Вопросы с длинными ответами (LA)
Главы
Лист
Цветок
Клетка – строение и функции
Пищеварительная система
Дыхательная система
Кровеносная система
Здоровье и гигиена
Среда обитания и адаптация
Курсы
Быстрые ссылки
Условия и политика
Условия и политика
2022 © Quality Tutorials Pvt Ltd.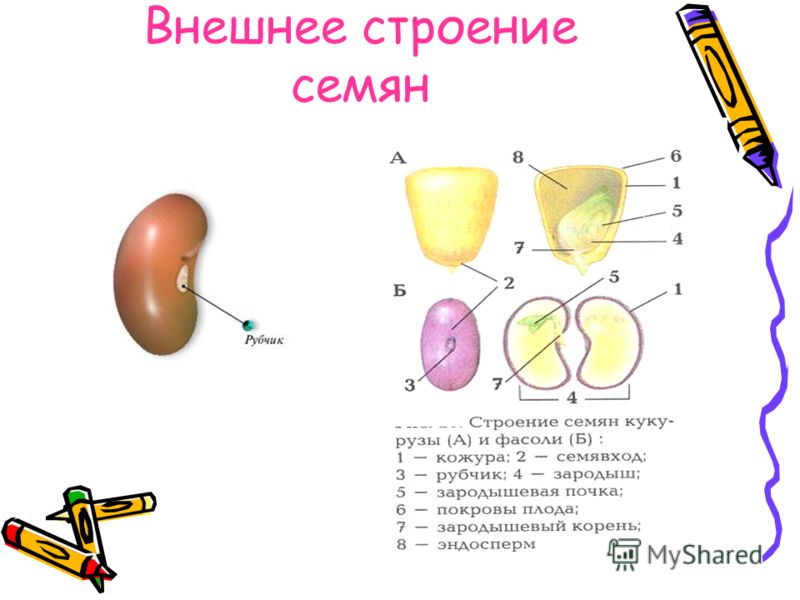 Все права защищены.
Список журналов
Все права защищены.
Список журналов
Книга арабидопсиса. 2008 г.; 6: e0119.
Опубликовано в Интернете 30 декабря 2008 г. doi: 10.1199/tab.0119
a, 1 и b, c
Информация об авторе Информация об авторских правах и лицензии Отказ от ответственности
Покой семян позволяет семенам преодолевать периоды, неблагоприятные для укоренившейся рассады, и поэтому важен для экологии растений и сельского хозяйства. Известно несколько процессов, участвующих в индукции покоя и в переключении из покоящегося состояния в прорастающее. В этой главе описывается роль растительных гормонов, различных вовлеченных тканей и генов, включая недавно идентифицированные гены в состоянии покоя и прорастания, а также использование анализа транскриптома, протеома и метаболома для изучения этих механистически недостаточно понятных процессов.
В этой главе описывается роль растительных гормонов, различных вовлеченных тканей и генов, включая недавно идентифицированные гены в состоянии покоя и прорастания, а также использование анализа транскриптома, протеома и метаболома для изучения этих механистически недостаточно понятных процессов.
Семя является важной стадией жизненного цикла высших растений в отношении их выживания как вида. Это дисперсионная единица растения, способная пережить период между созреванием семян и созданием следующего поколения в виде сеянцев после того, как они прорастут. Для этого выживания семя, в основном в сухом состоянии, хорошо подготовлено к тому, чтобы выдерживать длительные периоды неблагоприятных условий. Чтобы оптимизировать прорастание с течением времени, семя переходит в состояние покоя. Покой также препятствует прорастанию до сбора урожая. Были проведены многочисленные исследования, чтобы лучше понять, как прорастание контролируется различными факторами окружающей среды и применяемыми химическими веществами. Однако до сих пор очень мало известно о процессе, посредством которого зародыш выходит из семени для полного прорастания, и о том, как блокируется появление зародыша в покоящихся семенах (Bewley, 19).97).
Однако до сих пор очень мало известно о процессе, посредством которого зародыш выходит из семени для полного прорастания, и о том, как блокируется появление зародыша в покоящихся семенах (Bewley, 19).97).
Arabidopsis обладает периодом покоя семян, как и многие другие виды растений. Это свойство контролируется такими факторами окружающей среды, как свет, температура и время сухого хранения семян, а также генетическими факторами. Использование генетики и молекулярной генетики в арабидопсисе начинает проливать свет на некоторые аспекты механизма покоя и прорастания путем идентификации мутантов и генов, контролирующих эти процессы. В этом обзоре представлен обзор современных знаний о состоянии покоя и прорастания семян арабидопсиса, основанный главным образом на вкладе молекулярной генетики в изучение этого процесса, включая таблицу с генами, связанными с прорастанием/прорастанием. Несколько недавних обзоров (Finch Savage and Leubner-Metzger 2006, Finkelstein et al., 2008, Holdsworth et al. , 2008a) описывают больше деталей молекулярного механизма, которые были получены в основном из молекулярно-генетических исследований в сочетании с физиологическими экспериментами.
, 2008a) описывают больше деталей молекулярного механизма, которые были получены в основном из молекулярно-генетических исследований в сочетании с физиологическими экспериментами.
Развитие семян состоит из двух основных фаз: развития зародыша и созревания семян. Эмбриогенез, который является фазой морфогенеза, начинается с образования одноклеточной зиготы и заканчивается на стадии сердца, когда все структуры эмбриона сформированы (Mayer et al., 1991). За ней следует фаза роста, во время которой зародыш заполняет семенной мешок (Goldberg et al., 1994). В конце фазы роста эмбриона клеточное деление эмбриона прекращается (Raz et al., 2001). После этого семя, содержащее полноразмерный зародыш, подвергается созреванию, во время которого накапливаются пищевые запасы, развивается состояние покоя и устойчивость к высыханию (Goldberg et al., 19).94). Было опубликовано несколько подробных исследований, касающихся развития (Baud et al., 2002), метаболических (Fait et al., 2006), протеомных (Fu et al. , 2005; Gallardo et al., 2001, 2002; Rajjou et al., 2004 и Chibani et al., 2006, а экспрессия генов (Girke et al., 2000; Nakabayashi et al., 2005; Finch-Savage et al., 2007) изменяется во время развития семян, в сухих семенах и последующем прорастании
, 2005; Gallardo et al., 2001, 2002; Rajjou et al., 2004 и Chibani et al., 2006, а экспрессия генов (Girke et al., 2000; Nakabayashi et al., 2005; Finch-Savage et al., 2007) изменяется во время развития семян, в сухих семенах и последующем прорастании
Семя развитие широко изучалось с использованием мутантов, дефектных в различных аспектах этого процесса.Мутанты, затронутые в фазе морфогенеза, приводят к летальности или имеют фенотип проростка (Mayer et al., 1991; Мейнке, 1995). У мутантов прорастания семян нарушены свойства прорастания и покоя, что иногда сопровождается плейотропными эффектами, характерными для созревания, такими как непереносимость высыхания (Goldberg et al., 1994; Koornneef and Karssen, 1994).
Покой семян определяется как неспособность жизнеспособных семян прорастать в благоприятных условиях (Bewley, 1997; Finch-Savage and Leubner-Metzger, 2006). Покой у арабидопсиса следует охарактеризовать как физиологически неглубокий, что означает, что зародыши, освобожденные от окружающих структур, растут нормально, а покой теряется в результате влажного охлаждения (стратификации) или дозревания (Baskin, Baskin, 2004). Однако, в дополнение к слоям семенников и эндосперма, окружающим зародыш, потенциал роста зародыша также важен для преодоления ограничений этих структур и тем самым влияет на состояние покоя семени (Kucera et al., 2006).
Однако, в дополнение к слоям семенников и эндосперма, окружающим зародыш, потенциал роста зародыша также важен для преодоления ограничений этих структур и тем самым влияет на состояние покоя семени (Kucera et al., 2006).
Поскольку покой регулируется на разных фазах развития при взаимодействии с факторами окружающей среды, трудно определить момент установления генетических и физиологических различий. Эта трудность возникает потому, что все анализы состояния покоя основаны на прорастании семян, что является результатом баланса между степенью покоя и способностью зародыша преодолевать состояние покоя. Механически можно различать факторы, влияющие на покой и прорастание, на основе их влияния на прорастание, будь то ингибирующее или стимулирующее. Мутанты, которые прорастают лучше или быстрее, могут представлять гены, способствующие покою, или гены, подавляющие прорастание. Дальнейшее различие можно провести, определив время и место действия этих факторов (во время созревания или во время набухания семян, в зародыше, эндосперме или семеннике).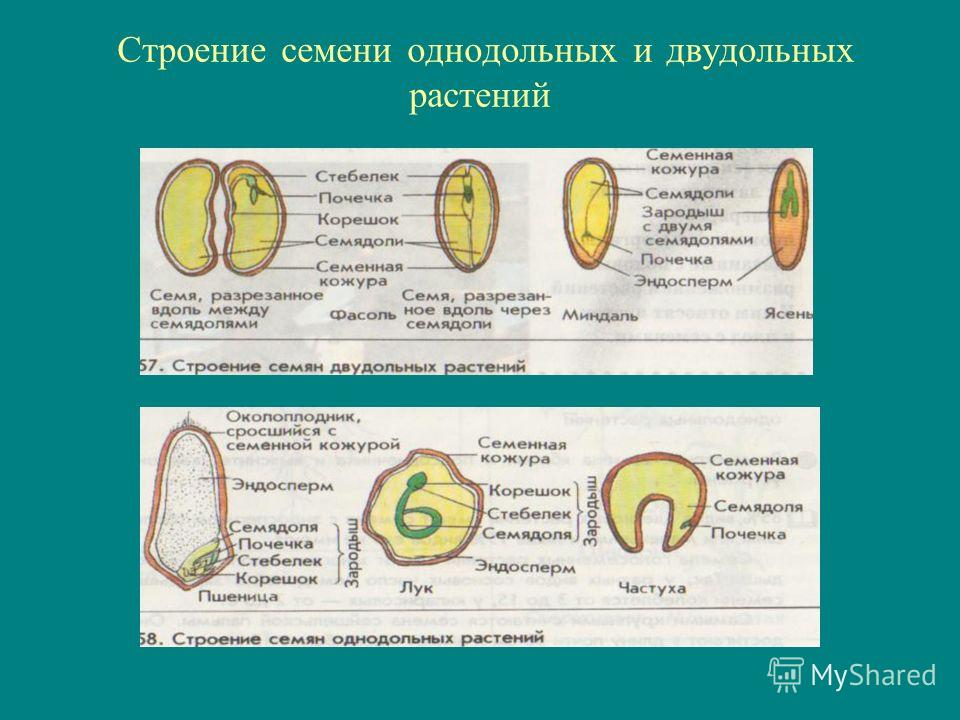 Взаимодействие между этими факторами и большое влияние окружающей среды как во время развития семян, так и во время набухания делают состояние покоя семян очень сложным признаком.
Взаимодействие между этими факторами и большое влияние окружающей среды как во время развития семян, так и во время набухания делают состояние покоя семян очень сложным признаком.
По определению, прорастание включает те события, которые начинаются с поглощения воды покоящимися сухими семенами и заканчиваются удлинением зародышевой оси (Bewley and Black, 1994). Поглощение воды семенами трехфазное; фаза I быстрое начальное поглощение; фаза II фаза плато, а в фазе III дальнейшее увеличение поглощения воды, однако, только при прорастании (Schopfer and Plachy, 1984; Bewley, 1997; Manz et al., 2005). Первыми признаками прорастания являются возобновление основных процессов, включая транскрипцию, трансляцию и репарацию ДНК с последующим удлинением клеток и, наконец, во время протрузии корешка возобновление клеточного деления (Barroco et al., 2005; Masubelele et al., 2005). Физически прорастание представляет собой двухстадийный процесс, при котором за разрывом тесты следует разрыв эндосперма. После разрыва микропилярного эндосперма образовавшимся корнем прорастание завершается (Karssen, 19).76; Хефер и Робертс, 1985 г.; Лейбнер-Мецгер и др., 1995; Крок и др., 2002; Петруцелли и др., 2003 г.; Лейбнер-Мецгер, 2003 г.; Лю и др., 2005). Анализы на прорастание арабидопсиса часто проводят на свету на свежесобранных или хранящихся в течение ограниченного времени семенах (Léon-Kloosterziel et al., 1996a). Другими параметрами являются скорость прорастания после различных периодов обработки холодом (Cutler et al., 1996) и прорастание в темноте (Meng et al., 2008; Kim et al., 2008). В дополнение к тестированию зрелых семян, проращивание незрелых семян, либо вырезанных из стручка, либо внутри плодов, отделенных от растения, может быть использовано для исследования генетической изменчивости на ранних стадиях развития семян (Raz et al., 2001).
После разрыва микропилярного эндосперма образовавшимся корнем прорастание завершается (Karssen, 19).76; Хефер и Робертс, 1985 г.; Лейбнер-Мецгер и др., 1995; Крок и др., 2002; Петруцелли и др., 2003 г.; Лейбнер-Мецгер, 2003 г.; Лю и др., 2005). Анализы на прорастание арабидопсиса часто проводят на свету на свежесобранных или хранящихся в течение ограниченного времени семенах (Léon-Kloosterziel et al., 1996a). Другими параметрами являются скорость прорастания после различных периодов обработки холодом (Cutler et al., 1996) и прорастание в темноте (Meng et al., 2008; Kim et al., 2008). В дополнение к тестированию зрелых семян, проращивание незрелых семян, либо вырезанных из стручка, либо внутри плодов, отделенных от растения, может быть использовано для исследования генетической изменчивости на ранних стадиях развития семян (Raz et al., 2001).
Открыть в отдельном окне
Проращивание семян арабидопсиса. Зрелое семя арабидопсиса может находиться как в состоянии покоя, так и в состоянии покоя. Спящее семя не прорастет, если оно будет подвергаться воздействию правильных условий окружающей среды (свет и вода). Покой семян может быть нарушен сухим хранением или холодным набуханием (стратификацией). Прорастание семян арабидопсиса происходит в два этапа: разрыв раковины с последующим разрывом эндосперма.
Спящее семя не прорастет, если оно будет подвергаться воздействию правильных условий окружающей среды (свет и вода). Покой семян может быть нарушен сухим хранением или холодным набуханием (стратификацией). Прорастание семян арабидопсиса происходит в два этапа: разрыв раковины с последующим разрывом эндосперма.
Поскольку ткани как материнского (теста), так и зиготного происхождения (эмбрион и эндосперм) способствуют прорастанию семян, генетический анализ покоя семян должен учитывать это различное происхождение тканей. Материнские эффекты, в отличие от зиготических факторов, наследуются по материнской линии и могут быть связаны с генетическим составом семенников, окружающих зародыш, но также могут быть связаны с генетическими различиями, связанными с факторами, которые переносятся в семя от материнского растения. О материнском наследовании можно судить по прорастанию семян, полученных в результате реципрокного скрещивания, где родительские генотипы используются как женские, так и мужские генотипы. Эндосперм – продукт оплодотворения. Однако геномный вклад самки-родителя в два раза больше, чем самца-родителя в этой триплоидной ткани, что в некоторых случаях может привести к различиям в реципрокных скрещиваниях, когда задействованы эффекты дозы. Недавно была показана роль этого единственного слоя эндосперма в состоянии покоя и необходимость ослабления этого клеточного слоя для прорастания (Müller et al., 2006; Bethke et al., 2007a; для обзора Holdsworth et al., 2008a).
Эндосперм – продукт оплодотворения. Однако геномный вклад самки-родителя в два раза больше, чем самца-родителя в этой триплоидной ткани, что в некоторых случаях может привести к различиям в реципрокных скрещиваниях, когда задействованы эффекты дозы. Недавно была показана роль этого единственного слоя эндосперма в состоянии покоя и необходимость ослабления этого клеточного слоя для прорастания (Müller et al., 2006; Bethke et al., 2007a; для обзора Holdsworth et al., 2008a).
Покой семян у Arabidopsis можно преодолеть с помощью факторов, способствующих прорастанию, таких как дозревание, свет, обработка холодом (также называемая стратификацией). Кроме того, применяемые химикаты, такие как гиббереллины и KNO 3 (Derkx and Karssen, 1993a; Alboresi et al. 2005; Ali-Rachedi et al. 2004), NO (Bethke et al., 2007b), также оказывают стимулирующее действие на прорастание (). Ни один из этих факторов окружающей среды не является абсолютным требованием для прорастания, потому что потребность в одном факторе зависит от других факторов, как показано Коне и Спруитом для взаимодействия между светом и температурой (19). 83). Эта потребность в экзогенных факторах очень сильно зависит от генотипа. Покой, высвобождаемый в результате сухого дозревания или воздействия холода или нитратов с последующим воздействием света, изучался у образца с островов Зеленого Мыса (Cvi). Оказалось, что чувствительность семян к холоду, нитратам и свету зависела от времени, в течение которого семена находились в сухом состоянии после дозревания. Семена стали сначала чувствительными к нитратам, затем к холоду и, наконец, к свету (Finch-Savage et al., 2007). Кроме того, было показано, что скорость нарастания чувствительности к сигналам внешней среды не была фиксированной, семена разных лет реагировали по-разному. Это согласуется с тем, что глубина покоя определяется не только генетически, но и окружающей средой во время формирования семян (Donohue, 2005).
Открыть в отдельном окне
Схематическое представление процессов, контролирующих состояние покоя и прорастание семян арабидопсиса. Семя арабидопсиса характеризуется зародышем с двумя семядолями и однослойным эндоспермом. Указаны стимулирующие прорастание (зеленые стрелки) и ингибирующие факторы (красные стрелки).
Семена, не находящиеся в состоянии покоя, которые некоторое время находились в неблагоприятных условиях для прорастания (например, набухшие семена, хранившиеся при относительно высокой температуре в темноте), могут снова войти в состояние покоя, которое называется вторичным состоянием покоя (Cone and Spruit, 1983; Деркс и Карссен, 1993b; Хилхорст, 2007).
Задача исследования покоя и прорастания состоит в том, чтобы определить природу важнейших регуляторов, которые предотвращают начало прорастания (покоя), запускают процесс прорастания и их взаимодействие. Кроме того, важно знать, как факторы окружающей среды, такие как свет и холод, влияют на эндогенные факторы, контролирующие прорастание.
Генетическая изменчивость может быть вызвана мутагенезом, но она также присутствует среди многих природных образцов (экотипов) арабидопсиса. Генетическая изменчивость прорастания может быть обнаружена при сравнении генотипов в идентичных условиях. Это означает, что не только условия испытания на всхожесть должны быть одинаковыми, но и условия роста при развитии семян и условия хранения, в том числе сроки хранения семян, должны быть одинаковыми. Кроме того, условия испытаний должны различать генотипы.
Тесты на всхожесть можно эффективно использовать для скрининга мутантов, поскольку можно анализировать большое количество. Однако изменчивость признака прорастания может привести к генетической неправильной классификации отдельных семян и, следовательно, к ложноположительным результатам при скрининге мутантов. Генотипы Arabidopsis Landsberg erecta (L er ) и Columbia (Col), которые в основном используются в исследованиях арабидопсиса, демонстрируют лишь низкий уровень покоя. Этот покой исчезает примерно через месяц после созревания (van der Schaar et al., 19).97). Из-за этого относительно низкого уровня покоя невозможно насытить мутации в генах покоя. Однако эту проблему можно решить, используя более спящие образцы (Koornneef et al. , 2000).
Генетические варианты, описанные в этом обзоре, объединяет то, что семена являются жизнеспособными, о чем свидетельствует их способность прорастать после специальной обработки, такой как разрушение семенной оболочки или применение определенных химикатов. Мутанты, которые не прорастают, потому что они смертельны, включая ранние мутанты семязачатка (Schneitz, 1999) и многие из так называемых летальных эмбрионов (Meinke, 1995) не описаны в этом обзоре, поскольку в основном они имеют дефекты развития, которые не контролируют специфически покой и прорастание.
Анализ естественной изменчивости
Arabidopsis является однолетним растением, для которого можно обнаружить большие различия в периоде покоя между образцами, собранными в природе (Ratcliffe, 1976). Куглер (1951) провел первое генетическое исследование различий в прорастании между образцами арабидопсиса. Различия в состоянии покоя поддаются генетическому анализу у арабидопсиса, как это было показано с помощью анализа локусов количественных признаков (QTL) для характеристик прорастания/покоя семян в нескольких популяциях рекомбинантных инбредных линий (RIL) (van der Schaar et al. , 19).97; Алонсо-Бланко и др., 2003 г. и Клеркс и др., 2004 г.). Эти популяции RIL были созданы путем скрещивания образцов, которые имеют разные уровни покоя семян, со стандартными лабораторными образцами L или . В анализе van der Schaar et al. (1997), L er /Col RIL выращивали в трех независимых экспериментах в теплицах и тестировали в различных условиях прорастания. Несмотря на небольшую разницу в состоянии покоя между родительскими линиями, всего можно было идентифицировать 14 QTL, большинство из которых оказывало лишь небольшое влияние. Многие из них были обнаружены только в определенных условиях прорастания или в семенах одного или двух сроков сбора урожая, что указывает на сложность генетического контроля различий в состоянии покоя.
Для популяции L er /Cvi RIL были описаны анализы QTL для состояния покоя семян, которые привели к идентификации локусов, влияющих на состояние покоя семян, что можно было подтвердить с помощью почти изогенных линий (NIL) (Alonso-Blanco et al. , 2003). Это исследование QTL выявило локусы, влияющие на потребность в послесозревании, измеренную как число дней сухого хранения семян, необходимое для достижения 50% всхожести (DSDS50). Таким образом, семь QTL были идентифицированы и названы Delay Of Germination 9.1166 ( СОБАКА ) 1–7. Для подтверждения и характеристики этих локусов были сконструированы NIL, несущие фрагменты интрогрессии Cvi на генетическом фоне L er . Анализ этих линий на прорастание подтвердил четыре QTL ( DOG1 , DOG2 , DOG3 и DOG6 ). Тот же тип анализов в популяции L er / Shahdara (Sha) RIL выявил четыре QTL. Три из них, DOG1 , DOG3 и DOG6 , находились в позициях, идентифицированных ранее (Alonso-Blanco et al., 2003) и, скорее всего, представляли одни и те же локусы. Кроме того, был идентифицирован QTL на хромосоме 2 (Clerkx et al., 2004).
Был клонирован первый из этих QTL покоя семян, DOG1 , основной QTL в популяции L er /Cvi (Bentsink et al. 2006). DOG1 экспрессируется во время развития семян, и транскрипт остается в зрелых сухих и после созревших семенах, но при впитывании транскрипт исчезает. DOG1 является членом небольшого семейства генов с неизвестной молекулярной функцией и ранее не был связан с состоянием покоя семян, что указывает на то, что естественная изменчивость может быть полезна для идентификации новых генов покоя семян (Bentsink et al., 2006).
Всего в настоящее время проанализировано семь популяций RIL, полученных от скрещиваний с L или , что привело к идентификации одиннадцати DOG QTL (Bentsink et al., неопубликовано). Этот скрининг потребности в дозревании достигает насыщения, поскольку в новых проанализированных популяциях те же самые QTL идентифицируются также в скрещивании, в котором L или не участвовали (X. Huang, M. Koornneef и K. Donohue, неопубликованные данные).
Анализы прорастания семян в ответ на холод и темноту показали возможность точного картирования и клонирования QTL, ответственных за устойчивые к холоду прорастания в темноте (Meng et al. , 2008). Авторы идентифицировали три основных локуса, ответственных за изменчивость этого признака в популяции Bay-0/Sha RIL. Один из этих QTL, CDG-1 ( Холодостойкая прорастание в темноте ) был локализован в той же области генома, что и локусы задержки прорастания DOG2 и DOG3 (Alonso-Blanco et al., 2003) и QTL, влияющие на контролируемая порча и скорость прорастания (Clerkx et al., 2004).
Другим признаком семян, для которого были описаны естественные вариации, является выделение слизи во время набухания. Образец Sha из Центральной Азии несет мутацию в гене MUM2 9.1166 (Macquet et al., 2007), что приводит к отсутствию слизи.
Гены и факторы, влияющие на покой/прорастание семян
Гены и/или факторы, индуцирующие покой семян или подавляющие прорастание
Первая важная стадия индукции покоя – вероятно, конец морфогенетической программы, когда все ткани присутствуют в зрелом зародыше сформировались, и эмбрион вступает в фазу остановки роста. АБК-НЕЧУВСТВИТЕЛЬНЫЙ3 ( ABI3 ), FUSCA3 ( FUS3 ) и LEAFY COTYLEDON ( LEC1 и LEC2 ) являются четырьмя ключевыми регуляторами, которые играют видную роль в контроле среднего и позднего развития семян (Meinke et al., 1994; Raz et al., 1994; Raz et al. , 2001). ABI3 , FUS3 и LEC2 кодируют родственные специфичные для растений факторы транскрипции, содержащие консервативный ДНК-связывающий домен B3 (Giraudat et al., 1992; Luerssen et al., 1998; Stone et al., 2001), тогда как ЛЭК1 кодирует субъединицу HAP3 фактора транскрипции, связывающего CCAAT (CBF, также известного как NF-Y (Lotan et al., 1998)). Все четыре мутанта abi3 , lec1 , lec2 и fus3 серьезно страдают при созревании семян и имеют некоторые общие фенотипы, такие как снижение периода покоя при созревании (Raz et al., 2001) и снижение экспрессии запаса семян. белки (Gutierrez et al. , 2007). Однако они также демонстрируют специфические фенотипы, такие как отсутствие деградации хлорофилла в сухих семенах (9).1165 abi3 ), снижение чувствительности к АБК ( abi3 и в меньшей степени lec1 ), накопление антоцианов ( fus3 , lec1 и, в меньшей степени, 16 lec2) , непереносимость к высыханию ( ABI3 , FUS3 и LEC1 ), или дефекты в идентичности семядолов ( LEC1 , FUS3 и LEC2 ) (Bäumble Et AT AT AT. ., 1994; Мейнке и др., 1994; Парси и др., 1994; Парси и Жиродат, 1997; Люерссен и др., 1998 г.; Виент и др., 2000; Раз и др., 2001; Стоун и др., 2001; Крой и др., 2003). Недавно было показано, что некоторые из фенотипов fus3 обусловлены плейотропными эффектами, вызванными укороченными генными продуктами мутантных аллелей. Прямые эффекты FUS3 , вероятно, ограничены состоянием покоя эмбрионов и определением идентичности клеток эпидермиса семядолей (Tiedemann et al. , 2008).
Ген LEC1 необходим для нормального развития на ранних и поздних фазах эмбриогенеза и достаточен для индукции эмбрионального развития в вегетативных клетках (Lotan et al., 1998). Потеря функции LEC1 приводит к прорастанию вырезанных зародышей на той же стадии (между 8–10 днями после опыления), что и у мутантов lec2 и fus3 , но раньше в процессе развития эмбриона, чем у мутантов abi3 (Raz и др., 2001). Десять субъединиц HAP3 (AHAP3) были идентифицированы у Arabidopsis, которые можно разделить на два класса на основе идентичности последовательностей в их центральном консервативном домене B (Kwong et al., 2003). LEC1 и близкородственная субъединица, LEC1-LIKE ( L1L ) составляют субъединиц AHAP3 типа LEC1 , тогда как остальные восемь обозначены как не относящиеся к типу LEC1 . Подобно LEC1 , L1L экспрессируется преимущественно во время развития семян. Однако супрессия экспрессии гена L1L индуцирует дефекты в развитии эмбрионов, которые отличаются от таковых у мутантов lec1 , указывая на то, что LEC1 и L1L играют разные роли в эмбриогенезе (Kwong et al., 2003).
LEC2 непосредственно контролирует программу транскрипции, участвующую в фазе созревания развития семян. Индукция активности LEC2 в проростках вызывает быстрое накопление РНК, обычно присутствующих в основном во время фазы созревания, включая запасные белки семян и липидные тела. Промоторы генов, кодирующих эти РНК созревания, все обладают мотивами RY (цис-элементы, связанные транскрипционными факторами домена B3) (Braybrook et al., 2006). Это дает убедительные доказательства того, что эти гены представляют собой мишени транскрипции ЛЭК2 . Одним из этих генов является DOG1 , первый ген покоя семян, ответственный за изменчивость, встречающуюся в природных популяциях, которая была идентифицирована на молекулярном уровне (Bentsink et al. , 2006).
Было показано, что ABI3 , FUS3 , LEC1 и LEC2 взаимодействуют как сеть для контроля различных аспектов созревания семян. Было показано, что LEC1 регулирует экспрессию как ABI3 , так и FUS3 (Kagaya et al., 2005), FUS3 и LEC2 частично избыточно контролируют экспрессию генов специфических для семян белков, а LEC2 локально регулирует экспрессию FUS3 в области семядолей (Kroj et al., 2003). Недавно было показано указание на избыточную регуляцию внутри этой группы генов (To et al., 2006). При анализе экспрессии ABI3 и FUS3 в различных мутантах с одинарным, двойным и тройным созреванием были идентифицированы множественные регуляторные связи между всеми четырьмя генами. Выяснилось, что одна из главных ролей LEC2 должен был повысить регулировку FUS3 и ABI3 . Мутация lec2 приводит к резкому снижению экспрессии ABI3 и FUS3 , и большинство фенотипов lec2 могут быть восстановлены конститутивной экспрессией ABI3 или FUS3 . Кроме того, было показано, что ABI3 и FUS3 положительно регулируют себя и друг друга, тем самым образуя петли обратной связи, необходимые для их устойчивой и единообразной экспрессии в эмбрионе. Наконец, LEC1 также положительно регулирует ABI3 и FUS3 в семядолях (To et al., 2006). Хотя среди этих четырех генов было идентифицировано несколько регуляторных связей, молекулярные механизмы, лежащие в основе этой сети, и нижестоящие мишени сети, связанные с индукцией покоя, все еще требуют дальнейшего изучения.
Помимо мутантов, влияющих на общее созревание семян, другие мутанты более конкретно влияют на покой семян, т.е. мутанты, которые изменяют биосинтез АБК или способ ее действия. АБК регулирует различные аспекты роста и развития растений, включая покой семян. Отсутствие АБК-индуцированного покоя позволяет семенам прорастать без гиббереллинов. Таким образом, отбор мутантов, прорастающих в присутствии ингибиторов биосинтеза ГА, таких как паклобутразол и тетциклацис, является эффективным способом выделения мутантов биосинтеза АБК (Léon-Kloosterziel et al. , 19).96б). Реципрокные скрещивания между диким типом и дефицитными по АБК мутантами aba1 показали, что покой контролируется генотипом АБК зародыша, а не материнского растения. Последнее является причиной относительно высоких уровней АБК, обнаруживаемых в семенах на полпути развития семян (Karssen et al., 1983). На этой фазе АБК может предотвращать преждевременное прорастание, о чем свидетельствуют материнские эффекты АБК у экстремальных двойных мутантов aba abi3-1 (Koornneef et al., 1989). Ключевыми генами биосинтеза АБК в процессе развития семян являются NCED6 и NCED9, оба являются членами семейства 9-цис-эпоксикаротиноидов диоксигенез (Lefebvre et al., 2006). В отличие от того, что было предложено Karssen et al (1983), теперь ясно показано, что эндогенная АБК необходима для поддержания покоя семян. У дикого типа уровни АБК снижаются в конце созревания семян и во время набухания за счет активности генов катаболизма АБК, принадлежащих к семейству P450 CYP707A (Okamoto et al. , 2006), что указывает на то, что уровни АБК могут изменяться на разных фазах прорастания семян. развития и прорастания со значительным влиянием на всхожесть. Кроме того, наблюдение, что ингибиторы биосинтеза АБК, такие как нор-флуразон, способствуют прорастанию (Debeaujon and Koornneef, 2000), показало, что поддержание покоя в набухших семенах является активным процессом, включающим синтез АБК de novo, что также было показано для покоящихся семян. партии образца Cvi (Ali-Rachedi et al., 2004).
АБК оказывает большое влияние на состояние покоя семян, поэтому можно ожидать, что нарушение передачи сигналов АБК также приводит к изменениям характеристик прорастания. Существенный прогресс был достигнут в характеристике таких путей передачи сигналов АБК (Bonetta and McCourt, 1998; Leung and Giraudat, 1998). Генетический скрининг для выявления сигнальных мутантов АБК был основан в первую очередь на ингибировании прорастания семян под действием АБК. Нечувствительные к АБК ( abi ) мутанты (Koornneef et al. , 1984) и несколько других, описанных позже (Rock, 2000 и Holdsworth et al., 2008a), способны прорастать и расти в присутствии концентраций АБК, ингибирующих рост дикого типа. Ожидалось, что такие скрининги дадут мутанты рецептора АБК и сигнальной трансдукции. Однако большинство генов рецепторов АБК были идентифицированы с использованием обратной генетики, при которой проводился скрининг характеристик прорастания, который часто не показывал или незначительно влиял на чувствительность АБК к прорастанию, а сам по себе покой часто не тестировался (обзор в Holdsworth et al., 2008a). Прямые скрининги мутантов, у которых прорастание семян предотвращается низкими концентрациями АБК, которые обычно позволяют прорастать семенам дикого типа, впервые были описаны Cutler et al. (1996), что привело к мутантам эра1 (усиленный ответ на АБК) в эра3 мутантов. Впоследствии с помощью аналогичных скринингов было выявлено множество дополнительных локусов, участвующих в удалении чувствительности к функции АБК, которые при мутации приводят к гиперчувствительности к АБК впитавшихся дозревших или влажно-охлажденных семян (Hugouvieux et al. , 2001; Xiong et al., 2001). ; Nishimura et al., 2004; Katagiri et al., 2005; Zhang et al., 2005; Pandey et al., 2006; Saez et al., 2006; Yoine et al., 2006; Nishimura et al., 2007) . Эти локусы кодируют разнообразные функции, в том числе связанные с трансляцией и метаболизмом РНК, путями деградации белков и фосфатазными компонентами сигнальных путей и факторов транскрипции (Holdsworth et al., 2008a). Судя по их влиянию на покой семян, эти два набора мутаций также изменяют регуляцию прорастания семян эндогенной АБК. 9Мутанты 1165 abi3 , abi4 и abi5 демонстрируют сниженную экспрессию различных генов созревания семян, но только мутанты abi3 не находятся в состоянии покоя, что совпадает с непереносимостью высыхания (Nambara et al., 1992, Ooms et al., 193). , Bies et al., 1999, Finkelstein et al., 2008) в сильных аллелях. Удивительно, но у мутантов abi4 и abi5 не наблюдалось состояния покоя или другого фенотипа созревания семян (Finkelstein, 1994; Finkelstein et al. , 2008), за исключением снижения некоторых мРНК, специфичных для созревания семян (Finkelstein and Lynch, 2000; Söderman, et al. др., 2000). Это может указывать на то, что другие гены дублируют функции этих специфичных для семян транскрипционных факторов, которые являются членами домена APETALA2 (9).1165 ABI4 , Финкельштейн и др., 1998; Söderman, et al., 2000) и семейство основных факторов лейциновой молнии ( ABI5 , Finkelstein and Lynch, 2000; Lopez-Molina et al., 2001).
Согласно недавнему отчету, уровни АБК могут позитивно регулироваться белком DELLA посредством усиления экспрессии XERICO (Zentella et al., 2007). Сверхэкспрессия XERICO , которая кодирует убиквитин лигазу E3 , приводит как к повышенному уровню АБК, так и к повышенной устойчивости к засухе, хотя механизм до сих пор неизвестен (Ko et al., 2006).
Класс мутантов, который был напрямую отобран на основе сниженного периода покоя, представляет собой мутанты rdo1-rdo4 (Léon-Kloosterziel et al. , 1996a; Peeters et al., 2002). Тот факт, что все четыре мутанта проявляют некоторые умеренные плейотропные эффекты у взрослых растений, указывает на то, что гены не специфичны для покоя/прорастания, но влияют и на другие процессы. Было показано, что RDO4 (переименованный в HUB1 ) кодирует белок C3HC4 Ring finger, участвующий в моноубиквитинировании гистона h3B, что указывает на роль моделирования хроматина в состоянии покоя семян (Liu et al., 2007). 9Мутант 1165 dag1-1 также проявляет пониженный покой, но в отличие от мутантов rdo этот эффект определяется материнским генотипом. Это согласуется с характером экспрессии гена DAG1 в сосудистой ткани развивающегося семени. DAG1 , который кодирует транскрипционный фактор DOF, может влиять на импорт соединений из материнского растения в семена (Papi et al., 2000). Это первый ген, идентифицированный как специфически участвующий в материнском контроле прорастания семян. Однако фенотип прорастания dag2 , мутант в родственном гене DAG2 , с характером экспрессии, аналогичным DAG1 , противоположен таковому у семян dag1 (Gualaberti et al. , 2002), показывая повышенный покой. Были выделены дополнительные мутанты со сниженным фенотипом покоя в других локусах, в том числе мутанты без явного плейотропного эффекта (Bentsink et al., 2006 и неопубликованные результаты M. Schwab и W. Soppe), что указывает на сложность генетической регуляции покоя семян. . Вместо отбора мутантов, которые прорастают, когда дикий тип все еще находится в состоянии покоя, Salaita et al. (2005) использовали скорость прорастания при 10°C в качестве критерия отбора при скрининге меченых активацией линий образца Col. Кроме двух tt мутантов (см. ниже), ни один из этих прорастающих при низких температурах ( ctg ) не был охарактеризован на молекулярном уровне.
Другая группа мутантов, у которых снижен период покоя семян, — это мутанты с измененной семенной оболочкой или кожурой (Debeaujon et al., 2000; обзор Debeaujon et al., 2007; Lepiniec et al., 2006). Семенная кожура является многофункциональным органом, который играет важную роль в питании зародыша во время развития семян и в последующей защите от вредных факторов окружающей среды (Debeaujon et al. , 2007). Семенная кожура образуется из двух покровов эпидермального происхождения, окружающих зрелую семяпочку. Развитие семенной кожуры из семязачатка было описано Beeckman et al. (2000).
Семенная кожура вместе со слоем эндосперма оказывает действие, ограничивающее прорастание, либо будучи непроницаемой для воды и/или кислорода, производя соединения, ингибирующие прорастание, либо благодаря своей механической устойчивости к выпячиванию корней. У арабидопсиса фенольные соединения и их производные, присутствующие во внутреннем слое тесты, называемом эндотелием, влияют на свойства семенной кожуры, влияющие на прорастание, о чем можно сделать вывод из сниженного фенотипа покоя многих мутантов тесты.
Мутанты семенной кожуры состоят из двух основных групп. Одна группа, пораженная флавоноидной пигментацией, представлена мутантами прозрачной тесты ( tt ) и прозрачной тесты glabra ( ttg ). Идентифицированы мутанты от tt1 до tt15 , ttg1 и ttg2 и banyuls ( ban ). Цвет мутантов tt варьирует от желтого до бледно-коричневого (Debeaujon et al., 2000). 9Мутанты 1165 Ban накапливают розовые флавоноидные пигменты в эндотелии незрелых семян, но не содержат проантоцианов из-за мутации в гене антоцианинредуктазы ( ANR ) (Xie et al., 2003), что приводит к появлению серовато-зеленых пятнистых зрелых семян. семена (Альберт и др., 1997; Девик и др., 1999). У мутанта ttg1 отсутствует слизь и трихомы, и он нарушен в морфологии внешнего слоя семенной кожуры, а также в продукции пигмента. Многие из мутантов семенной кожуры уже клонированы, для получения более подробной информации см. дополнительную таблицу 1 онлайн и Debeaujon et al. (2007).
Вторая группа представлена мутантами, пораженными в строении тесты. Мутант с аберрантной формой тесты ( ats ), мутантный в гене KANADI 4 (McAbee et al., 2006), производит один покров вместо двух покровов, наблюдаемых у семязачатков дикого типа, и демонстрирует меньший период покоя.
Гены и/или факторы, уменьшающие период покоя семян или повышающие способность семян к прорастанию
Прорастание семян арабидопсиса находится под фотоконтролем с помощью фитохромов. Поэтому следует ожидать, что мутанты с дефицитом фитохрома будут затронуты прорастанием семян. Сложность системы фитохромов обусловлена наличием различных типов фитохромов, для которых пять генов в геноме арабидопсиса кодируют разные, но родственные апопротеины (Sharrock and Quail, 19).89). Кроме того, на прорастание могут влиять различные способы действия фитохрома, описанные как реакция на очень низкую плотность энергии (VLFR), реакция на низкую плотность энергии (LFR) и реакция на высокую освещенность (HIR), которые имеют свою собственную зависимость от плотности энергии (рассмотрено Касаль и Санчес, 1998). Мутанты, лишенные фитохрома B (phyB), демонстрируют пониженную чувствительность к красному свету, что указывает на то, что phyB играет основную роль в прорастании семян. PhyA может индуцировать прорастание только после длительного набухания семян (Shinomura et al. , 19).94). Подробные спектры действия на прорастание семян, полученные у мутантов дикого типа, phyA и phyB , выявили типичную красно-/дальнекрасную (R/FR)-обратимую LFR, опосредованную phyB, тогда как реакция прорастания, опосредованная phyA, оказалась VLFR с чувствительностью к свету в 10 4 раз выше (Shinomura et al, 1996). Наблюдение, что двойные мутанты phyA phyB демонстрируют некоторое светозависимое прорастание, указывает на участие другой R/FR-обратимой фоторецепторной системы (Yang et al., 19).95; Poppe and Schäfer, 1997), возможно, опосредованное phyC, D и/или E.
Хотя основная роль фитохрома заключается в индуцированной светом стимуляции прорастания семян, его роль в наступлении покоя или установлении потребности в свете на это указывают эксперименты Маккалоу и Шропшира (1970) и Хейса и Кляйна (1974). Эти авторы показали, что отношение R/FR, испытываемое материнским растением и, следовательно, во время созревания семян, влияет на последующее поведение зрелых семян при прорастании. Мунир и др. (2001) показали, что условия фотопериода во время формирования семян также могут влиять на прорастание семян. Однако этот эффект сильно зависел от генотипа. Кроме того, оказывается, что пути, опосредованные фитохромами, необходимы для выхода из состояния покоя, индуцированного холодом (Donohue et al., 2007). Низкие температуры во время созревания семян вызывали покой семян, который не мог быть преодолен в течение 9 лет.1165 hy2-1 (дефицит фитохромного хромофора, общего для всех пяти фитохромов) мутант.
Растительный гормон гиббереллин играет важную роль в стимулировании прорастания семян. Мутанты с дефицитом GA не могут прорастать без экзогенных GA (Koornneef and van der Veen, 1980; Mitchum et al., 2006). Биосинтез ГК de novo необходим во время набухания, как было сделано из наблюдения, что ингибиторы биосинтеза ГА, такие как паклобутразол и тетциклацис, предотвращают прорастание (Karssen et al., 19).89), если АВА отсутствует. Как и ожидалось, также мутанты, передающие сигнал GA, такие как мутант sleepy1 ( sly1 ), который был отобран в ходе скрининга супрессоров нечувствительного к АБК мутанта abi1-1 (Steber et al. , 1998), и гены, кодирующие белок Della, такие как RGL2 (Lee et al., 2002; Bassel et al., 2004) имеют фенотипы с дефектом прорастания. Для мутантов рецепторов GA ( gid1a , gid1b , gid1c ) необходимо было сконструировать тройные мутанты, чтобы увидеть этот фенотип из-за избыточности функции этих генов (Griffith et al., 2006, Iuchi et al., 2007 г., обзор Хирано и др., 2008 г.). ГК могут способствовать прорастанию за счет своей способности преодолевать ограничения прорастания, которые существуют у семян, требующих дозревания, света и холода. Это привело к предположению, что такие факторы окружающей среды могут индуцировать биосинтез ГК на ранних фазах прорастания. В настоящее время хорошо документированы изменения содержания GA и экспрессии генов биосинтеза и катаболизма GA во время выхода из покоя и прорастания (Yamauchi et al., 2007).
Эффект фитохрома (света) был подтвержден Yamaguchi et al. (1998), которые показали, что один из двух ферментов 3-β-гидроксилазы, кодируемый геном GA4H , индуцируется фитохромом в прорастающих семенах. Механизм сигнальных белков GA и эффект света в настоящее время хорошо установлен. Важнейшими в этом являются белки DELLA, которые подавляют действие GA, RGL2 (репрессор GA1-3, подобный 2) является основным DELLA, регулирующим прорастание семян. Эти белки DELLA расщепляются протеосомой 26S, поэтому заряженные GA белки GID1 DELLA взаимодействуют с белком F-бокса SLY1, необходимым для убиквитинирования DELLA (Steber, 2007, Finkelstein et al., 2008). Важным компонентом сигнальной трансдукции индуцированного светом прорастания является фактор транскрипции bHLH 9.1165 PIF1 ( Phytochrome Interacting Factor 1 ), также называемый PIL5 ( Phytochrome-Interacting factor 3-Like 5 ) (Oh et al., 2004, 2006, Castillon et al., 2007), который может связываться с Pfr, что впоследствии приводит к деградации протеосом (Oh et al., 2006). Это увеличивает уровни GA, потому что гены биосинтеза GA, такие как GA3ox1 и GA3ox2, репрессируются с помощью PIF1 / PIL5 , тогда как катаболический ген GA GA2ox и гены, кодирующие белки DELLA, активируются (Oh et al. , 2007). Эти более высокие уровни GA дополнительно приводят к инактивации белков DELLA, как описано выше. Somnus ( SOM ), который кодирует локализованный в ядре белок цинкового пальца CCCH-типа, является другим геном, действующим ниже по течению от PIL5 (Kim et al., 2008). Мутант сом прорастает в темноте независимо от различных световых режимов. Ким и др. (2008) показали, что PIL5 активирует экспрессию SOM путем связывания непосредственно с промотором SOM . Предполагается, что PIL5 регулирует метаболические гены ABA и GA частично через SOM.
Обработка холодом также стимулирует биосинтез GA (Yamauchi et al., 2004) путем индукции генов GA3ox1 и GA3ox2. Этот холодовой эффект опосредуется светостабильным транскрипционным фактором bHLH SPATULA (SPT), который подавляет экспрессию этих генов (Penfield et al., 2005). Кроме того, холод может повышать чувствительность семян к ГА, поскольку он также влияет на мутанты с дефицитом ГА (Debeaujon and Koornneef, 2000). Тот факт, что часто стратификация оказывается более эффективной, чем обработка ГА (Alonso-Blanco et al., 2003), свидетельствует о том, что стратификация влияет и на другие факторы, способствующие прорастанию.
Брассиностероиды (BRs), природные растительные стероидные гормоны, обнаруживаемые в большом количестве видов растений (Clouse and Sasse, 1998), также участвуют в контроле прорастания арабидопсиса. Сигнал BR приводит к снижению чувствительности к АБК и тем самым стимулирует прорастание, хотя нормальное прорастание BR-дефицитных и BR-сигнальных мутантов указывает на отсутствие абсолютной потребности BR для прорастания (Steber and McCourt, 2001). БР могли преодолеть отсутствие всхожести sleepy1 мутант, вероятно, путем обхода его потребности в GA через независимый от GA механизм (Steber and McCourt, 2001; Finkelstein et al., 2008).
Мутанты в передаче сигналов этилена также подвержены влиянию на их реакцию прорастания. Этилен вырабатывается в следовых количествах почти всеми высшими растениями и участвует в контроле процессов роста и развития, которые варьируются от прорастания до старения. Часто семена, реагирующие на этилен, чувствительны к свету для прорастания (Kepczynski and Kepczynska, 19).97). Нечувствительные к этилену мутанты, такие как etr и ein2 , прорастают хуже или после более длительного периода дозревания, чем дикий тип (Bleecker et al., 1988; Beaudoin et al., 2000). Мутанты ein2 и etr гиперчувствительны к АБК (Beaudoin et al., 2000; Ghassemian et al., 2000), что согласуется с наблюдением, что мутанты ein2 были выделены как супрессоры abi1-1 . Среди мутантов, отобранных в качестве энхансеров нечувствительного к АБК мутанта 9, появился мутант ctr1 , характеризующийся конститутивным ответом на этилен.Моногенные мутанты 1165 abi1-1 и ctr1 также слабо устойчивы к АБК (Beaudoin et al., 2000). Эти наблюдения в сочетании с фенотипом отсутствия покоя у двойного мутанта ein2 abi3-4 показали, что этилен отрицательно регулирует состояние покоя семян путем ингибирования действия АБК (Beaudoin et al. , 2000). Однако Ghassemian et al., (2000) и Chiwocha et al. (2005) показали, что, помимо передачи сигналов, немного более спящие мутанты, нечувствительные к этилену, такие как ein2 и etr1-2 имеют более высокие уровни АБК. Более высокие уровни гормонов, стимулирующих прорастание, указывают на компенсационные эффекты. Присутствие перекрестных помех между передачей сигналов сахара и этиленом было подтверждено нечувствительным к сахару фенотипом ctr1 (Gibson et al., 2001) и гиперчувствительным к сахару фенотипом etr (Zhou et al., 1998). По-видимому, сигнализация АБК, этилена и сахара сильно взаимодействует на уровне прорастания и раннего роста проростков.
Известно, что ауксины играют важную роль в эмбриогенезе. Однако его роль в регуляции прорастания и укоренения проростков оставалась неясной (Kucera et al., 2006). Сам по себе ауксин обычно не считался важным для контроля прорастания семян, но предполагалось, что взаимодействие между ауксином, АБК, ГК и этиленом влияет как на прорастание, так и на укоренение проростков (Fu and Harberd, 2003; Ogawa et al. , 2003; Chiwocha et al., 2005, Carrera et al., 2008; Liu, PP et al., 2007, рассмотрено Holdsworth et al., 2008a). Анализ экспрессии репортера ауксина DR5:GUS показал, что ауксин накапливается во время эмбриогенеза и присутствует в семени после набухания. Экспрессия наблюдалась на кончике корешка перед прорастанием в одном исследовании (Liu, PP et al., 2007) и по всему эмбриону в конце эмбриогенеза в другом исследовании (Ni et al., 2001). Анализ экспрессии транскриптома показал, что РНК, кодирующая переносчики ауксина AUX1, PIN2 и PIN7, сильно активировалась в ответ на лечение ga1 мутантных семян с GA (Ogawa et al., 2003), и что транспортеры как оттока, так и притока активируются в дозревших семенах по сравнению с покоящимися семенами (Carrera et al., 2008). Это может указывать на роль этих транспортеров в прорастании как таковом или в установлении верхушки корня и гравитропизма после появления корешка. Более четкие генетические доказательства роли ауксина в прорастании были получены при анализе регуляции Auxin Response Factor10 ( ARF10 ) с помощью микроРНК (миРНК) miR160 (Liu, PP et al. , 2007). miRNAs, как было показано, подавляют гены-мишени на посттранскрипционном уровне и играют решающую роль в широком диапазоне онтогенетических процессов (Dugas & Bartel, 2004). Было показано, что трансгенные семена, экспрессирующие устойчивую к miR160 форму ARF10 ( mARF10 ), были гиперчувствительны к ингибированию прорастания экзогенной АБК, тогда как эктопическая экспрессия miR160 приводила к снижению чувствительности к АБК (Liu, PP et al., 2007). ).
Эти результаты указывают на роль ауксина в путях, связанных с прорастанием, и предполагают, что взаимодействия между ауксином и сигнальными путями АБК могут способствовать способности семян к прорастанию. Анализ функции ключевых компонентов передачи сигналов ауксина в отношении после созревания, потенциала прорастания и силы роста может выявить новые роли ауксина в этих процессах.
Соединения, которые были идентифицированы как важные стимуляторы прорастания, в основном с использованием фармакологических инструментов, представляют собой несколько азотсодержащих соединений, в том числе газообразный оксид азота (NO) (Bethke et al. , 2006), нитрит (NO 2 − ) и нитрат (NO 3 − ) (Alboresi et al., 2005; Bethke et al., 2007a для обзора). Предполагается (Bethke et al., 2007a), что все соединения азота влияют на прорастание посредством преобразования в NO. Ферментативное производство NO происходит в основном с помощью нитратредуктазы как побочного продукта катаболизма липидов или синтазы оксида азота (Crawford and Guo, 2005). Также было продемонстрировано неферментативное превращение нитрита в NO, и было высказано предположение, что оно имеет особое значение для семян (Bethke et al., 2007a). Наблюдение Alboresi et al. (2005) тот факт, что высокие уровни нитратов, которые накапливаются у мутантов с дефицитом нитратредуктазы, приводят к уменьшению периода покоя, подразумевает, что нитратредуктаза не является существенной либо потому, что NO генерируется этим неферментативным путем, либо нитрат действует сам по себе.
Влияние NO заключается в том, что он может действовать как антиоксидант (Lamatinna et al. , 2003). Однако также сообщается, что NO ингибирует каталазу, что приводит к более высокому уровню H 2 O 2 и активным формам кислорода (АФК), что оказывает противоположное действие антиоксидантов (Bethke et al., 2007a). АФК являются побочным продуктом β-окисления запасенных жирных кислот семян. Тем самым увеличиваются АФК, что само по себе может облегчать состояние покоя (Bailly et al., 2004). Однако также были предложены другие и дополнительные механизмы, включающие взаимодействие с ферментами катаболизма АБК и сигналами света и ГК (Finkelstein et al., 2008, Bethke et al., 2007b)
Семена могут сохраняться в течение длительного времени без прорастания, как при хранении в сухих условиях, так и при закапывании в почву. Эти семена образуют банк семян, ожидая, пока условия окружающей среды не станут благоприятными для прорастания. Несмотря на сильную устойчивость многих семян к высыханию, хранение также в сухих условиях в конечном итоге приводит к потере всхожести. Было высказано предположение, что такие соединения, как определенные сахара и белки, такие как LEA (обильный поздний эмбриогенез) (Skriver and Mundy, 1990) и белки теплового шока (Hsps) (Wehmeyer and Vierling, 2000; Hong and Vierling, 2001), которые накапливаются на более поздних стадиях развития семян, выполняют защитную функцию от высыхания. Однако мутанты, такие как abi5 , которые показывают сильное снижение некоторых белков LEA (Finkelstein and Lynch, 2000), не имеют явных дефектов при хранении семян. Мутант Hsp101 с нокаутом демонстрирует потерю термотолерантности во время прорастания семян (Hong and Vierling, 2001), но в оптимальных условиях прорастание кажется незатронутым, указывая на то, что, по крайней мере, Hsp101 не играет общей функции в выживании высушенных семян. Экспрессия этих белков теплового шока в процессе развития контролируется специфичным для семян фактором транскрипции теплового стресса HsfA9., который регулируется белком ABI3 (Kotak et al., 2007).
Присутствие генетической изменчивости в отношении способности к хранению очевидно для генотипов, не переносящих высыхание, таких как мутанты abi3 , lec1 , fus3 и dog1 (Meinke et al. , 1994; ., 2006), а также сообщается о воздействии на мутанты тесты (Debeaujon et al., 2000). Анализ QTL в линиях L er /Cvi и L er /Sha RIL (Bentsink et al., 2000; Clerkx et al., 2004) также выявил генетическую изменчивость по этому признаку, которая, однако, не была связана с уровнями олигосахаридов ряда рафинозы, сегрегирующих в одной и той же популяции.
Для семени важно, чтобы после периода, в течение которого оно не могло прорасти, оно могло вырасти в сильнорослый саженец, способный конкурировать с другими саженцами. Факторы, которые важны для укоренения рассады, вероятно, в основном связаны с наличием и мобилизацией материалов для хранения. Накопление запасных белков и липидов нарушено у мутантов созревания семян, таких как lec1 , lec2 и fus3 , которые, хотя и прорастают на фильтровальной бумаге, не могут быть эффективно перенесены в почву из этого субстрата, в отличие от других семян. , и требуют укоренения в качестве рассады на среде с добавлением сахара, прежде чем их можно будет пересадить в почву. wrinkled1 ( wri1 ; Focks and Benning, 1998) мутант специфически воздействует на соединения для хранения семян с сильным снижением содержания масла в семенах. Кроме того, он демонстрирует пониженный потенциал прорастания и пониженное укоренение проростков и иначе, чем дикий тип, реагирует на сигналы сахара и АБК (Cernac et al., 2006). WRI1 кодирует транскрипционный фактор семейства AP2/EREB (Cernac and Benning, 2004), который сам регулируется LEC2 (Baud et al., 2007). WRI1 контролирует углеродный обмен в развивающихся семенах и является необходимым условием для накопления масла в семенах.
Мобилизация запасов пищи во время прорастания и раннего роста рассады требует активности ферментов, расщепляющих липиды и углеводы. Расщепление запасных липидов представляет собой сложный процесс, включающий ряд стадий, для которых многие гены были идентифицированы в последние годы, и их влияние на мутации в этих генах было изучено у мутантов и двойных мутантов (обзор в Penfield et al. , 2007; Грэм и др., 2008). Жирные кислоты запасаются в масляных телах в виде триацилглицеринов (ТАГ), которые гидролизуются липазами. Высвобожденные жирные кислоты передаются глиоксисомам (пероксисомам, в которых происходит глиоксилатный цикл) через переносчик ABC, кодируемый белком CTS (comatose) (Russell et al., 2000; Footitt et al., 2002). Здесь происходит β-окисление, превращающее жирные кислоты, активированные в сложные эфиры ацил-КоА, в ацетил-КоА, который затем превращается в четырехуглеродные сахара. Эти сахара затем транспортируются в митохондрии, откуда они превращаются в малат и транспортируются в цитозоль для глюконеогенеза или используются для дыхания. Профилирование экспрессии генов показало, что флавоноиды отсутствовали в cts и kat2 –1 мутантные семена, но накапливались в присутствии сахарозы. Это указывает на существенную роль β-окисления в индукции генов биосинтеза флавоноидов. Тот же анализ показал, что CTS функционирует очень поздно в фазе II прорастания и что его функция необходима для экспрессии специфических наборов генов, связанных с важным биохимическим путем, связанным с укоренением и выживанием проростков (Carrera et al. , 2007).
Белки, которые играют роль в β-окислении, представляют собой LACS (ацил-КоА-синтетаза), ACX (ацил-КоА-оксидаза), MFP2 (многофункциональный белок 2) и KAT2 (3-кето-ацил-КоА-тиолаза). Глиоксилатный цикл включает цитратсинтазу (CSY), изоцитратлиазу (ICL), малатсинтазу (MLS) и фосфоенолпируваткарбоксикиназу (PCK1) (обзор Penfield et al., 2007; Graham et al., 2008).
Как и ожидалось от мутантов с дефицитом запасных липидов, многие мутанты в этих генах, описанных выше (или двойные мутанты, когда дуплицированные гены контролируют этап) имеют дефекты удлинения гипокотилей в темноте и укоренения проростков, которые могут быть устранены сахарами (Eastmond et al. и др., 2000; Корна и др., 2004). Следовательно, глиоксилатный цикл можно охарактеризовать как несущественный для прорастания арабидопсиса, хотя он играет важную физиологическую роль в укоренении проростков. Однако подмножество одиночных мутантов по мобилизации липидов ( cts , kat2 ) и двойные (в случае acx и csy ) демонстрируют сильно сниженную всхожесть, что объясняет, почему мутанты cts были выделены при скрининге семян на всхожесть (Russell et al. , 2000). Общим для всех этих мутантов является то, что они действуют в глиоксисоме и участвуют в транспорте в (CTS) или экспорте из (CSY, Pracharoenwattana et al., 2005, Graham, 2007 для обзора) или в β-окислении в этой органелле. Это говорит о том, что продукт(ы) β-окисления, по-видимому, обладает функцией, способствующей прорастанию. Природа этого сигнала, которая не связана с отсутствием мобилизации липидов как таковой, неизвестна. Пенфилд и др. (2007) упоминают ряд возможностей, таких как биосинтез жасмоната или окислительно-восстановительный контроль. Однако прямое участие этих факторов маловероятно, поскольку другие мутанты по этим факторам не обнаруживают фенотипа прорастания (Penfield et al., 2007).
Примером другого гена, контролирующего метаболизм в развивающихся и прорастающих семенах, является пластидная пируваткиназа ( PKP1 ), контролирующая последнюю стадию гликолиза. Мутанты Pkp1 , у которых снижено накопление масла, содержат меньше хлорофилла и токоферола и, следовательно, дефект приживаемости проростков (Andre et al. , 2007; Baud et al., 2007).
Сахара задерживают раннюю рассаду. Было показано, что этот процесс тесно связан с биосинтезом и передачей сигналов растительных гормонов, в частности, с АБК (обзор см. в Finkelstein and Gibson, 2001; Gazzarini and McCourt, 2001; León and Sheen, 2003; Rook et al., 2006; Деккерс и Смикенс, 2007; Роньони и др., 2007). Мутанты, нечувствительные к ингибирующему действию глюкозы и сахарозы на раннее развитие проростков, были выделены несколькими группами и идентифицированы мутанты с дефицитом АБК (т.е. aba2 / isi4 / gin1 / sis4 and aba3 / gin5 ) and ABA insensitive4 ( abi4 / sun6 / isi3 / gin6 / sis5 ) как нечувствительные к сахару (Arenas-Huertero et al., 2000; Huijser et al., 2000; Laby et al., 2000; Rook et al., 2001). Тот факт, что ABI4 кодирует домен AP2, содержащий фактор транскрипции, который связывает CE1-подобный элемент, присутствующий во многих промоторах, регулируемых ABA и сахаром (Finkelstein et al. , 19).98; Ниу и др., 2002; Acevedo-Hernandez et al., 2005) связали регуляцию сахара с передачей сигналов ABA. Однако есть еще много примеров процессов и генов, которые регулируются АБК и сахаром. Недавно Деккерс и соавт. (2008) показали, что ABI3 является ключевым регулятором нечувствительной к глюкозе ( GIN ) передачи сигналов, что согласуется с предыдущими отчетами, показывающими, что сверхэкспрессия ABI3 (либо AtABI3 , либо CnABI3 ) у арабидопсиса приводит к сверхчувствительный фенотип (Finkelstein et al., 2002; Zeng and Kermode, 2004). Те же авторы показали, что ABA и передача сигналов глюкозы перекрываются в большей степени, чем первоначально предполагалось (Dekkers et al., 2008). Недавно было установлено, что DOG1 также участвует в опосредованном АБК сигнальном пути сахара у проростков. Эта роль специфична для аллеля GSQ5 / DOG1 Cvi, поскольку добавление глюкозы индуцировало экспрессию аллеля GSQ5 / DOG1 Cvi, тогда как аллель L er и Col не реагировали (Teng et al. , 2008).
В нескольких недавних исследованиях была проанализирована экспрессия генома в семенах арабидопсиса как на уровне транскриптома, так и на уровне протеома (обзор Holdsworth et al., 2008b). Эти исследования выявили важную роль трансляции в прорастании и выходе из состояния покоя. Раджоу и др. (2004) сообщили, что транскрипция не является ограничивающим этапом завершения прорастания после впитывания семян, не находящихся в состоянии покоя. Набухание семян в присутствии избытка -аманитина (специфического ингибитора ДНК-зависимой РНК-полимеразы II) не ингибировало завершение прорастания, тогда как ингибитор трансляции циклогексамид полностью ингибировал прорастание. Это указывает на важность хранящейся в сухих семенах мРНК, которая используется для трансляции в набухших семенах (Rajjou et al., 2004)
Несколько недавних исследований показали, что важные изменения транскрипции и белка происходят в «сухих» семенах во время хранения и что эти изменения могут быть направлены на выход из состояния покоя по мере созревания семян. Эти исследования показывают, что может происходить как накопление специфических белков (Chibani et al., 2006), так и генных транскриптов (Bove et al., 2005, Leubner-Metzger, 2005). «Сухое» состояние, очень четко описанное Holdsworth et al. (2008b), представляют собой зрелые семена, содержащие 5–10% воды в зависимости от вида. Эта остаточная вода неравномерно распределена в тканях семени, что указывает на то, что некоторые области (гидратированные карманы внутри клеток) могут содержать достаточно воды для поддержания экспрессии генов (Leubner-Metzger, 2005).
Транскриптом
Фазы развития во время позднего эмбриогенеза и последующего прорастания характеризуются пространственными и временными паттернами экспрессии генов (Nambara et al., 2000; Parcy et al., 1997). Было проведено несколько исследований, чтобы понять процессы, происходящие в «сухом» семени, например, при дозревании. Каррера и др. (2008) провели исследование, чтобы понять взаимосвязь между АБК и дозреванием. Было показано, что мутанты, дефектные по синтезу АБК в семенах ( aba1-1 ) или восприятия ( abi1-1 ), которые не проявляют покоя, демонстрируют изменения глобальной экспрессии генов в результате сухого дозревания, которые были сравнимы с изменениями, происходящими в семенах дикого типа. Таким образом, действие АБК было отделено от экспрессии генов, регулируемых дозреванием и сухим хранением. Авторы также используют свою специфичную для семян онтологию генов (GO) TAGGIT, которая была описана в более ранней статье (Carrera et al., 2007), которая облегчает идентификацию и визуализацию сигнатуры прорастания.
Считается, что виды мРНК, хранящиеся в сухих семенах, необходимы для позднего эмбриогенеза и прорастания семян. Чтобы дифференцировать эти классы, Nakabayaski et al. (2005) проанализировали профили экспрессии, полученные из семян через 6, 12 и 24 часа после набухания. Было показано, что эмбриональные гены в значительной степени подавляются в течение 6 часов после набухания и что индукция метаболических генов, участвующих в прорастании, начинается примерно через 12 часов после набухания. В том же исследовании было обнаружено, что расположенные вверх по течению области генов, которые высоко экспрессируются в сухих семенах, в большей степени представляют ABRE (элементы, реагирующие на АБК). ABRE требует второго элемента для формирования функционального комплекса ответа ABA. Соединительный элемент (CE) является таким мотивом и вызывает ответ на ABA, когда он расположен рядом с ABRE (ABRC; Shen and Ho., 19).95). Действительно было показано, что гены, содержащие как ABRE, так и CE, экспрессируются в семенах на более высоких уровнях. В дополнение к этому мотив RY/Sph (известно, что он регулирует специфичную для семян экспрессию; Bäumlein et al., 1992; Hattori et al., 1992) проявлял обогащение семян при ассоциации с ABRE (Nakabayashi et al., 2005).
Пенфилд и др. (2006) установили относительную роль зародыша и эндосперма в контроле прорастания семян и укоренения проростков. Они показали, что эндосперм реагирует как на АБК, так и на гиббереллины, но что АБК, в частности, регулирует ядерную, а не кодируемую пластидами экспрессию генов фотосинтеза у эмбрионов. Они также показали, что Экспрессия ABI4 ограничена эмбрионом, что объясняет основные различия в ответе эмбрионов на ABA и определяет роль ABI4 как репрессора распада липидов. Более того, экспрессия ABI5 в эндосперме определяет вторую область измененной передачи сигналов ABA в микропилярном колпачке эндосперма. Наконец, сигнальные мутанты АБК зародыша и эндосперма демонстрируют пространственную специфичность действия АБК при прорастании семян. Был сделан вывод, что одноклеточный эндосперм играет активную роль в регуляции прорастания семян арабидопсиса (Penfield et al., 2006).
В других исследованиях были получены данные микрочипов, относящиеся к прорастанию в различных условиях. Эксперименты с использованием GA-дефицитных семян (Ogawa et al., 2003) описали временные изменения количества транскриптов в GA-дефицитных мутантных семенах, стимулированных к прорастанию обработкой GA, или в непрорастающих контрольных образцах, напитанных водой. У образца Cvi арабидопсиса изучали цикличность покоя в различных состояниях покоя и после созревания. Данные подтверждают механизм баланса гормонов ABA-GA, контролирующий цикличность через состояния покоя, который зависит от синтетических и катаболических путей обоих гормонов (Cadman et al. , 2006). Эти же авторы изучали влияние дозревания, низкой температуры, нитратов и света на экспрессию генов при выходе из покоя (Finch-Savage et al., 2007). Здесь они показывают, что наборы генов покоя и дозревания количественно реагируют на специфические сигналы окружающей среды. Недавно Бассель и соавт. (2008) применили химико-генетический подход для выявления малых молекул, подавляющих прорастание. Авторы идентифицировали три механистически различных ингибитора, которые они впоследствии использовали для выделения транскрипционных сигнатур, определяющих прорастание семян. Данные сравнивали с данными, опубликованными Carrera et al. (2007).
Чтобы помочь визуализировать данные, упомянутые выше, браузер экспрессии мРНК семян был разработан с использованием платформы электронных флуоресцентных пиктограмм (eFP). Этот eFP входит в состав пакета онлайн-инструментов биоинформатики Bio-Array (BAR) (www.bar.utoronto.ca). Начальный браузер eFP, разработанный и описанный Basssel et al. (2008) визуализирует экспериментальные данные экспрессии генов, нанесенные на пиктографические изображения тканей и условий, из которых были получены образцы РНК. TAGGIT GO (Carrera et al., 2007) также был интегрирован в браузер eFP.
Протеом
Было проведено несколько анализов протеома для анализа содержания белка в семенах арабидопсиса в зависимости от состояния покоя и прорастания с использованием двумерных гелей. Галлардо и др. (2001, 2002a) описали содержание белка, связанное с прорастанием и праймированием семян (обработка перед прорастанием, которая преодолевает период покоя семян), а также роль гиббереллинов в прорастании семян. Кроме того, была показана важность биосинтеза метионина для прорастания семян арабидопсиса и роста проростков (Gallardo et al., 2002b). Результаты показали, что метионинсинтаза и саденозилметионинсинтетаза являются основными компонентами, контролирующими метаболизм при переходе от покоящегося к высокоактивному состоянию во время прорастания семян.
Недавно был опубликован анализ протеомики в масштабе генома, в котором динамика протеома была описана для различных органов растений с использованием масс-спектрометрии (Baerenfaller et al. , 2008). Это исследование включает данные о дозревших «сухих» семенах, в этих семенах был обнаружен 13 901 различных пептидов, представляющих 3 789 белков. Всего было идентифицировано 133 биомаркера, специфичных для семян. Эти биомаркеры были обогащены определенными функциональными классами, а именно: эмбриональное развитие, заканчивающееся периодом покоя семян (GO:0009793), связывание липидов (GO:0019915), транспорт липидов (GO:0006869), покой семян (GO:0010162) и реакцию на воду (GO:0009415). Хотя в этой статье описывается только исследование белка в «сухих» семенах после созревания, карта специфических пептидов семян показывает, что протеомику можно использовать в качестве рутинного метода оценки, сравнивающего различные состояния покоя и прорастания.
Метаболом
Fait et al. (2006) проанализировали метаболические профили с помощью газовой хроматографии-масс-спектрометрии (ГС-МС) семян на протяжении всего развития и прорастания. Авторы заключают, что созревание семян связано со значительным снижением содержания большинства сахаров, органических кислот и аминокислот, что свидетельствует об их включении в запасы. Кроме того, оказалось, что переход от накопления запасов к высыханию семян был связан с основным метаболическим переключением, приводящим к накоплению различных сахаров, органических кислот, богатых азотом аминокислот и метаболитов, полученных из шикимата. В отличие от этого, стратификация семян была связана со снижением содержания нескольких метаболических промежуточных продуктов, накопленных во время высыхания семян, что означает, что эти промежуточные продукты могут поддерживать метаболическую реорганизацию, необходимую для прорастания семян. Одновременно уровни других метаболитов значительно увеличились во время стратификации и еще больше увеличились во время прорастания sensu stricto, что подразумевает их важность для прорастания и укоренения проростков. Метаболические переключения во время созревания и прорастания семян также были связаны с различными паттернами экспрессии генов, кодирующих продукты генов, связанных с метаболизмом, что было определено с помощью полуколичественной ОТ-ПЦР и анализа общедоступных данных микрочипов. В заключение, это исследование дает исчерпывающую картину скоординированных изменений в первичном метаболизме, лежащих в основе развития и прорастания семян у арабидопсиса. Кроме того, они подразумевают, что метаболическая подготовка к прорастанию и эффективному укоренению проростков начинается уже во время высыхания семян и продолжается дополнительными отчетливыми метаболическими переключениями во время стратификации и раннего прорастания (Fait et al., 2006).
Покой и прорастание — очень сложные признаки, находящиеся под контролем большого количества генов. В последние годы число вовлеченных мутантов и генов быстро увеличилось. Эти исследования подтвердили решающую роль растительных гормонов ГК и АБК. Однако он также показал участие и комплексное взаимодействие с другими растительными гормонами, такими как этилен, брассиностероиды и ауксин. Кроме того, были обнаружены новые игроки, такие как генов HUB , влияющих на модификации гистонов, и гены, такие как DOG1 , для которых мутанты с потерей функции показали свою важность, но для которых биохимическая функция неизвестна. Использование транскриптомного и протеомного анализа уточняет изменчивость и сходство путей, на которые влияет покой и прорастание, и обеспечивает основу для изучения роли отдельных генов в этих путях. В будущих исследованиях, помимо разграничения покоя и стимулирования прорастания, также будут приниматься во внимание временные и пространственные компоненты, а также конкретные роли и механизмы восприятия факторов окружающей среды как во время развития семян, так и во время хранения семян и прорастания семян. Вместе это должно позволить глубже проникнуть в суть этой важной и интригующей биологической системы.
Образец цитирования: Бенцинк Л. и Корнниф М. (2008) Покой семян и прорастание. Книга об арабидопсисе 6:e0119. doi:10.1199/tab.0119
elocation-id: e0119
† Эта глава была обновлена 31 декабря 2008 г. Первоначальная версия была заархивирована и доступна в формате PDF по адресу http://dx.doi. орг/10.1199/таб.0050.
- Albert S. , Delseny M., Devic M. BANYULS, новый негативный регулятор биосинтеза флавоноидов в семенной оболочке арабидопсиса. Завод Ж. 1997;116(1):289–299. [PubMed] [Google Scholar]
- Альборези А., Гестин К., Лейдекер М. Т., Беду М., Мейер К., Труонг Х. Н. Нитрат, сигнал, облегчающий состояние покоя семян арабидопсиса. Окружающая среда растительной клетки. 2005; 286(1):500–512. [PubMed] [Google Scholar]
- Ali-Rachedi S., Bouinot D., Wagner M.H., Bonnet M., Sotta B., Grappin P., Jullien M. Изменения уровней эндогенной абсцизовой кислоты во время выхода из покоя и поддержания зрелости семена: исследования с экотипом островов Зеленого Мыса, покоящейся моделью Arabidopsis thaliana. Планта. 2004;2196(1):479–488. [PubMed] [Google Scholar]
- Alonso-Blanco C., Bentsink L., Hanhart C.J., Blankestijn-De Vries H., Koornneef M. Анализ естественной аллельной изменчивости в локусах покоя семян Arabidopsis thaliana. Генетика. 2003;1646(1):711–729. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Алонсо Дж. , Хираяма Т., Роман Г., Нуризаде С., Экер Дж. Р. EIN2, бифункциональный преобразователь этилена и реакции на стресс у арабидопсиса. Наука. 1999;2846(1):2148–2155. [PubMed] [Академия Google]
- Андре С., Фрёлих Дж. Э., Молл М. Р., Беннинг С. Гетеромерный пластидный комплекс пируваткиназы, участвующий в биосинтезе масла семян арабидопсиса. Растительная клетка. 2007;196(1):2006–2022. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Аренас-Уэртеро Ф., Арройо А., Чжоу Л., Шин Дж., Леон П. Анализ нечувствительных к глюкозе мутантов арабидопсиса, gin5 и gin6, показывает центральную роль растительного гормона АБК в регуляции вегетативного развития растений сахаром. Гены Дев. 2000; 146(1):2085–209.6. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Беренфаллер К., Гроссманн Дж., Гробей М.А., Халл Р., Хирш-Хоффманн М., Яловски С., Циммерманн П., Гроссниклаус У., Груиссем В., Багинский С. Протеомика в масштабе генома раскрывает модели генов Arabidopsis thaliana и динамику протеома. Наука. 2008;3206(1):938–941. [PubMed] [Google Scholar]
- Bailly C. Активные формы кислорода и антиоксиданты в биологии семян. Семенная наука. Рез. 2004;146(1):93–107. [Google Scholar]
- Барроко Р. М., Ван П. К., Бергервоет Дж. Х., Де В. Л., Грут С. П., Инзе Д., Энглер Г. Роль механизмов клеточного цикла в возобновлении постэмбрионального развития. Завод Физиол. 2005;1376(1):127–140. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Баскин Дж. М., Баскин К. С. Система классификации покоя семян. Семенная наука. Рез. 2004;146(1):1–16. [Google Scholar]
- Basssel G. W., Zielinska E., Mullen R. T., Bewley J. D. Понижающая регуляция генов DELLA не является существенной для прорастания семян томата, сои и арабидопсиса. Завод Физиол. 2004;1366(1):2782–2789. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Бассель Г. В., Фунг П., Фриман Чоу Т-ф, Фунг Дж. А., Проварт Н. Дж., Катлер С. Р. Выяснение программы транскрипции прорастания с использованием малых молекул. Завод Физиол. 2008;1476(1):143–155. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Бод С., Бутин Дж. П., Микель М., Лепинец Л., Роша К. Комплексный обзор развития семян в экотипе Arabidopsis thaliana Ws. Завод Физиол. Биохим. 2002;406(1):151–160. [Google Scholar]
- Baud S., Mendoza M.S., To A., Harscoet E., Lepiniec L., Dubreucq B. WRINKLED1 определяет регуляторное действие LEAFY COTYLEDON2 на метаболизм жирных кислот во время созревания семян у арабидопсиса. Плант Дж. 2007; 506 (1): 825–838. [PubMed] [Google Scholar]
- Baumlein H., Nagy I., Villarroel R., Inze D., Wobus U. Цис-анализ промотора гена семенного белка: консервативный повтор RY CATGCATG внутри блока легумина необходим для тканеспецифическая экспрессия гена легумина. Завод Ж. 1992;26(1):233–239. [PubMed] [Google Scholar]
- Баумляйн Х., Мизера С., Луерсен Х., Кёлле К., Хорстманн С., Вобус У., Мюллер А. Дж. Ген FUS3 Arabidopsis thaliana является регулятором экспрессии генов на позднем эмбриогенезе. . Плант Дж. 1994;66(1):379–387. [Google Scholar]
- Бодуан Н., Серизе С., Гости Ф., Жиродат Дж. Взаимодействие между абсцизовой кислотой и сигнальными каскадами этилена. Растительная клетка. 2000;126(1):1103–1115. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Бекман Т., Де Райке Р., Виан Р., Инзе Д. Гистологическое исследование развития семенной кожуры у Arabidopsis thaliana. Дж. Плант Рез. 2000;1136(1):139–148. [Google Scholar]
- Bennett M.J., Marchant A., Green H.G., May S.T., Ward S.P., Millner P.A., Walker A.R., Schulz B., Feldmann K.A. Ген AUX1 арабидопсиса: пермеазоподобный регулятор корневого гравитропизма. Наука. 1996;2736(1):948–950. [PubMed] [Google Scholar]
- Бенцинк Л., Алонсо-Бланко К., Вройгденхил Д., Тесньер К.Дж.Ю., Грут С.П.С., Корннеф М. Генетический анализ растворимых в семенах олигосахаридов в отношении способности семян арабидопсиса храниться. Завод Физиол. 2000;1246(1):1595–1604. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Бенцинк Л. , Джоуэтт Дж., Ханхарт С.Дж., Корнниф М. Клонирование DOG1, локуса количественных признаков, контролирующего покой семян у арабидопсиса. проц. Натл. акад. науч. США. 2006;1036(1):17042–17047. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Бетке П. К., Либурель И. Г., Джонс Р. Л. Оксид азота снижает период покоя семян у арабидопсиса. Дж. Эксп. Бот. 2006;576(1):517–526. [PubMed] [Google Scholar]
- Бетке П. К., Либурель И. Г., Джонс Р. Л. Оксид азота в состоянии покоя и прорастания семян. В: Брэдфорд К., Ноногаки Х., редакторы. Развитие семян, покой и прорастание. 1. Том. 6. Оксфорд, Великобритания: издательство Blackwell Publishing; 2007а. стр. 153–171. [Академия Google]
- Bethke P.C., Libourel I.G., Aoyama N., Chung Y.Y., Still D.W., Jones R.L. Алейроновый слой Arabidopsis реагирует на оксид азота, гиббереллин и абсцизовую кислоту и является достаточным и необходимым для покоя семян. Завод Физиол. 2007b; 1436(1):1173–1188. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Bewley J. D., Black M. Seeds; Физиология развития и прорастания. Нью-Йорк: Пленум Пресс; 1994. [Google Scholar]
- Бьюли Дж. Д. Прорастание семян и покой. Растительная клетка. 1997;96(1):1055–1066. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Bies N., Silva Conceicao A., Giraudat J., Koornneef M., Leon-Kloosterziel K.M., Valon C., Delseny M. Важность домена B2 Белок Arabidopsis ABI3 для регуляции генов EM и 2S альбумина. Завод Мол. биол. 1999;406(1):1045–1054. [PubMed] [Google Scholar]
- Bleecker AB, Estelle MA, Somerville C., Kende H. Нечувствительность к этилену, обусловленная доминантной мутацией в Arabidopsis thaliana. Наука. 1988;2416(1):1086–1089. [PubMed] [Google Scholar]
- Бонетта Д., Маккорт П. Генетический анализ путей передачи сигнала АБК. Тенденции Растениевод. 1998;36(1):231–235. [Google Scholar]
- Bove J., Lucas P., Godin B., Oge L., Jullien M., Grappin P. Анализ экспрессии генов с помощью кДНК-AFLP выявляет ряд новых сигнальных сетей и трансляционный контроль во время нарушения покоя семян. у Nicotiana plumbaginifolia. Завод Мол. биол. 2005;576(1):593–612. [PubMed] [Академия Google]
- Braybrook S.A., Stone S.L., Park S., Bui A.Q., Le B.H., Fischer R.L., Goldberg R.B., Harada J.J. Гены, напрямую регулируемые LEAFY COTYLEDON2, дают представление о контроле созревания эмбриона и соматического эмбриогенеза. проц. Натл. акад. науч. США. 2006;1036(1):3468–3473. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Cadman C.S., Toorop PE, Hilhorst HW, Finch-Savage WE. Профили экспрессии генов семян Arabidopsis Cvi во время цикла покоя указывают на общий основной механизм контроля покоя. Плант Дж. 2006; 466 (1): 805–822. [PubMed] [Академия Google]
- Carrera E., Holman T., Medhurst A., Peer W., Schmuths H., Footitt S., Theodoulou F.L., Holdsworth M.J. Профилирование экспрессии генов выявляет определенные функции АТФ-связывающего кассетного переносчика COMATOSE в конце фазы II прорастание. Завод Физиол. 2007;1436(1):1669–1679. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Каррера Э. , Холман Т., Медхерст А., Дитрих Д., Футит С., Теодулу Ф. Л., Холдсворт М. Дж. Дозревание семян — это дискретный путь развития, связанный с специфические генные сети арабидопсиса. Плант Дж. 2008; 536 (1): 214–224. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Casal JJ, Sánchez R.A. Фитохромы и прорастание семян. Семенная наука. Рез. 1998;86(1):317–329. [Google Scholar]
- Castillon A., Shen H., Huq E. Факторы взаимодействия фитохромов: центральные игроки в опосредованных фитохромами сетях передачи световых сигналов. Тенденции Растениевод. 2007;126(1):514–521. [PubMed] [Google Scholar]
- Cernac A., Benning C. WRINKLED1 кодирует белок домена AP2/EREB, участвующий в контроле биосинтеза запасных соединений у арабидопсиса. Плант Дж. 2004; 406 (1): 575–585. [PubMed] [Академия Google]
- Сернак А., Андре К., Хоффманн-Беннинг С., Беннинг К. WRI1 требуется для прорастания семян и укоренения рассады. Завод Физиол. 2006;1416(1):745–757. [Статья бесплатно PMC] [PubMed] [Google Scholar]
- Chang C. , Kwok S.F., Bleecker A.B., Meyerowitz E.M. Ген ETR1, отвечающий за реакцию на этилен Arabidopsis: сходство продукта с двухкомпонентными регуляторами. Наука. 1993;2626(1):539–544. [PubMed] [Google Scholar]
- Чибани К., Ли-Рачеди С., Джоб С., Джоб Д., Жюльен М., Граппин П. Протеомный анализ покоя семян арабидопсиса. Завод Физиол. 2006;1426(1):1493–1510. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Чивоча С. Д. С., Катлер А. Дж., Абрамс С. Р., Амброуз С. Дж., Ян Дж., Росс А. Р. С., Кермод А. Р. Мутация etr1-2 в Arabidopsis thaliana влияет на абсцизовую кислоту, ауксин, пути метаболизма цитокинина и гиббереллина при поддержании покоя семян, влажностном охлаждении и прорастании. Плант Дж. 2005; 426 (1): 35–48. [PubMed] [Google Scholar]
- Clerkx E.J.M., El Lithy M.E., Vierling E., Ruys G.J., Blankestijin-De Vries H., Groot S.P.C., Vreugdenhil D., Koornneef M. Анализ естественной аллельной изменчивости прорастания семян арабидопсиса и признаков долголетия семян между образцами Landsberg erecta и Shakdara с использованием популяции новой рекомбинантной инбредной линии. Завод Физиол. 2004;1356(1):432–443. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Clouse S., Sasse J.M. Брассиностероиды: основные регуляторы роста и развития растений. Анну. Преподобный Завод. Физиол. Завод Мол. биол. 1998;496(1):427–451. [PubMed] [Google Scholar]
- Cone JW, Spruit CJP. Условия впитывания и покой семян Arabidopsis thaliana. Физиол. Завод. 1983;596(1):416–420. [Google Scholar]
- Коне Дж. В., Кендрик Р. Э. Кривые плотности потока и спектры действия для стимулирования и ингибирования прорастания семян у мутантов арабидопсиса thaliana L. Planta дикого типа и длинного гипокотиля. 1985;1636(1):43–54. [PubMed] [Google Scholar]
- Корна Дж. Э., Жермен В., Уорд Дж. Л., Бил М. Х., Смит С. М. Использование липидов, глюконеогенез и рост проростков у мутантов арабидопсиса, лишенных фермента глиоксилатного цикла малатсинтазы. Дж. Биол. хим. 2004;2796(1):42916–42923. [PubMed] [Google Scholar]
- Кроуфорд Н. М., Го Ф. К. Новое понимание метаболизма оксида азота и регуляторных функций. Тенденции Растениевод. 2005;106(1):195–200. [PubMed] [Google Scholar]
- Катлер С., Гассемиан М., Бонетта Д., Куни С., Маккорт П. Протеин-фарнезилтрансфераза, участвующая в передаче сигнала АБК у арабидопсиса. Наука. 1996;2736(1):1239–1241. [PubMed] [Google Scholar]
- Dall’Osto L., Cazzaniga S., North H., Marion-Poll A., Bassi R. Мутант Arabidopsis aba4-1 обнаруживает специфическую функцию неоксантина в защите от фотоокислительного стресс. Растительная клетка. 2007;196(1):1048–1064. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Debaujon I., Leon-Kloosterziel KM, Koornneef M. Влияние тесты на покой семян, прорастание и продолжительность жизни у Arabidopsis thaliana. Завод Физиол. 2000;1226(1):403–414. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Debaujon I., Koornneef M. Потребность в гиббереллине для прорастания семян Arabidopsis thaliana определяется как характеристиками тесты, так и эмбриональной АБК. Завод Физиол. 2000;1226(1):415–424. [Статья бесплатно PMC] [PubMed] [Google Scholar]
- Debaujon I., Peeters A.J.M., Leon-Kloosterziel K.M., Koornneef M. Ген TRANSPARENT TESTA 12 арабидопсиса кодирует мультилекарственный вторичный транспортер-подобный белок, необходимый для секвестрации флавоноидов в вакуолях. эндотелия семенной кожуры. Растительная клетка. 2001;136(1):853–871. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Дебожон И., Лепинец Л., Пурсель Л., Рутабул Ж-М. Развитие семенной кожуры и период покоя. В: Брэдфорд К., Ноногаки Х., редакторы. Развитие семян, покой и прорастание. 1. Том. 6. Оксфорд, Великобритания: издательство Blackwell Publishing; 2007. С. 25–43. [Google Scholar]
- Деккерс Б. Дж., Смикенс С. Сахар и абсцизовая кислота регулируют прорастание и переход к росту проростков. В: Брэдфорд К., Ноногаки Х., редакторы. Развитие семян, покой и прорастание. 1. Том. 6. Оксфорд, Великобритания: издательство Blackwell Publishing; 2007. стр. 305–322. [Академия Google]
- Деккерс Б. Дж., Шуурманс Дж. А., Смикенс С. С. Взаимодействие между сигналами сахара и абсцизовой кислоты во время раннего развития проростков арабидопсиса. Завод Мол. биол. 2008;676(1):151–167. [Статья бесплатно PMC] [PubMed] [Google Scholar]
- Derkx M.P.M., Karssen C.M. Изменчивость потребности семян Arabidopsis thaliana в свете, гиббереллине и нитратах в зависимости от времени сбора урожая и условий сухого хранения. J. Физиол растений. 1993а; 1416(1):574–582. [Google Scholar]
- Derkx M.P.M., Karssen C.M. Изменение чувствительности к свету и нитратам, но не к гиббереллинам, регулирует сезонный режим покоя семян Sisymbrium officinale. Окружающая среда растительной клетки. 1993б;166(1):469–479. [Google Scholar]
- Devic M., Guilleminot J., Debaujon I., Bechtold N., Bensaude E., Koornneef M., Pelletier G., Delseny M. Ген BANYULS кодирует DFR-подобный белок и является маркером раннего развития семенной кожуры. Плант Дж. 1999;196(1):387–398. [PubMed] [Google Scholar]
- Донохью К. , Дорн Л., Гриффит С., Ким Э., Агилера А., Полисетти С. Р., Шмитт Дж. Влияние окружающей среды и генетики на прорастание Arabidopsis thaliana в полевых условиях. Эволюция. 2005;596(1):740–757. [PubMed] [Google Scholar]
- Донохью К., Хешель М.С., Чианг Г.К., Батлер С.М., Баруа Д. Фитохром опосредует реакцию прорастания на несколько сезонных сигналов. Окружающая среда растительной клетки. 2007;306(1):202–212. [PubMed] [Google Scholar]
- Дугас Д. В., Бартель Б. Регуляция микроРНК экспрессии генов у растений. Курс. мнение биол. растений 2004;76(1):512–520. [PubMed] [Google Scholar]
- Dugas DV, Bartel B. Индукция сахарозой miR398 арабидопсиса репрессирует две супероксиддисмутазы Cu/Zn. Завод Мол. биол. 2008;676(1):403–417. [PubMed] [Академия Google]
- Истмонд П. Дж., Жермен В., Ланга П. Р., Брайс Дж. Х., Смит С. М., Грэм И. А. Постгерминативный рост и катаболизм липидов в семенах масличных культур, лишенных глиоксилатного цикла. проц. Натл. акад. науч. США. 2000;976(1):5669–5674. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Истмонд П. Дж., Грэм И. А. Пересмотр роли глиоксилатного цикла в семенах масличных культур. Тенденции Растениевод. 2001;66(1):72–77. [PubMed] [Google Scholar]
- Eastmond P. J. SUGAR-DEPENDENT1 кодирует триацилглицероллипазу с пататиновым доменом, которая инициирует расщепление запасного масла в прорастающих семенах арабидопсиса. Растительная клетка. 2006;186(1):665–675. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Fait A., Angelovici R., Less H., Ohad I., Urbanczyk-Wochniak E., Fernie A.R., Galili G. Развитие и прорастание семян арабидопсиса связаны с различными во времени метаболическими переключателями. Завод Физиол. 2006;1426(1):839–854. [Статья бесплатно PMC] [PubMed] [Google Scholar]
- Feinbaum RL, Ausubel FM. Транскрипционная регуляция гена халконсинтазы Arabidopsis. Мол. Клетка. биол. 1988;86(1):1985–1992. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Finch-Savage WE, Leubner-Metzger G. Покой семян и контроль прорастания. Новый Фитол. 2006;1716(1):501–523. [PubMed] [Академия Google]
- Finch-Savage W.E., Cadman C.S., Toorop P.E., Lynn J.R., Hilhorst H.W. Высвобождение семян из состояния покоя у Arabidopsis Cvi путем сухого дозревания, низкой температуры, нитратов и света демонстрирует общие количественные закономерности экспрессии генов, управляемые специфическими для окружающей среды ощущениями. Плант Дж. 2007; 516 (1): 60–78. [PubMed] [Google Scholar]
- Финкельштейн Р., Ван М. Л., Линч Т. Дж., Рао С., Гудман Х. М. Локус реакции абсцизовой кислоты арабидопсиса ABI4 кодирует доменный белок APETALA2. Растительная клетка. 1998;106(1):1043–1054. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Финкельштейн Р., Ривз В., Ариизуми Т., Стебер К. Молекулярные аспекты покоя семян. Анну. Преподобный завод биол. 2008;596(1):387–415. [PubMed] [Google Scholar]
- Финкельштейн Р. Р. Мутации в двух новых локусах ABA-ответа арабидопсиса аналогичны мутациям abi3. Плант Дж. 1994; 56 (1): 765–771. [Google Scholar]
- Финкельштейн Р. Р., Линч Т. Дж. Ген реакции на абсцизовую кислоту арабидопсиса ABI5 кодирует основной фактор транскрипции лейциновой молнии. Растительная клетка. 2000;126(1):599–609. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Финкельштейн Р. Р., Гибсон С. И. Взаимодействия ABA и сахара, регулирующие развитие: перекрестные разговоры или голоса в толпе. Курс. мнение биол. растений 2001;56(1):26–32. [PubMed] [Google Scholar]
- Финкельштейн Р. Р., Гампала С. С., Рок С. Д. Передача сигналов абсцизовой кислотой в семенах и проростках. Растительная клетка. 2002; 146 (Приложение) (1): S15–S45. [Статья бесплатно PMC] [PubMed] [Google Scholar]
- Focks N., Benning C. Wrinkled1: Новый мутант арабидопсиса с низким содержанием масла в семенах с дефицитом специфичной для семян регуляции углеводного обмена. Завод Физиол. 1998;1186(1):91–101. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Focks N. , Sagasser M., Weisshaar B., Benning C. Характеристика tt15, нового прозрачного мутанта раковины Arabidopsis thaliana (L.) Heynh. Планта. 1999;2086(1):352–357. [PubMed] [Google Scholar]
- Футит С., Слокомб С. П., Ларнер В., Куруп С., Ву Ю., Ларсон Т., Грэм И., Бейкер А., Холдсворт М. Контроль прорастания и мобилизации липидов с помощью COMATOSE, арабидопсис, гомолог ALDP человека. EMBO Дж. 2002; 216 (1): 2912–2922. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Fu Q., Wang B.C., Jin X., Li HB, Han P., Wei K.H., Zhang X.M., Zhu Y.X. Протеомный анализ и обширная идентификация белка из сухих, прорастающих Семена арабидопсиса и молодые саженцы. Дж. Биохим. Мол. биол. 2005;386(1):650–660. [PubMed] [Google Scholar]
- Fu X. D., Harberd N. P. Ауксин способствует росту корней арабидопсиса, модулируя реакцию гиббереллина. Природа. 2003;4216(1):740–743. [PubMed] [Академия Google]
- Gallardo K., Job C., Groot S.P.C., Puype M., Demol H., Vandekerck-hove J., Job D. Протеомный анализ прорастания и затравки семян арабидопсиса. Завод Физиол. 2001;1266(1):835–848. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Gallardo K., Job C., Groot SPC, Puype M., Demol H., Vandekerck-hove J., Job D. Важность биосинтеза метионина для прорастания семян арабидопсиса и рост рассады. Физиол. Завод. 2002;1166(1):238–247. [PubMed] [Академия Google]
- Gallardo K., Job C., Groot S.P.C., Puype M., Demol H., Vandekerck-hove J., Job D. Протеомика прорастания семян арабидопсиса. Сравнительное исследование семян дикого типа и семян с дефицитом гиббереллина. Завод Физиол. 2002;1296(1):823–837. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Газзаррини С., Маккорт П. Генетические взаимодействия между сигнальными путями ABA, этилена и сахара. Курс. мнение биол. растений 2001;46(1):387–391. [PubMed] [Google Scholar]
- Жермен В., Рилотт Э. Л., Ларсон Т. Р., Шерсон С. М., Бехтольд Н., Карде Дж. П., Брайс Дж. Х., Грэм И. А., Смит С. М. Потребность в 3-кетоацил-КоА-тиолазе-2 при развитии пероксисом , бета-окисление жирных кислот и расщепление триацилглицерина в липидных телах проростков арабидопсиса. Плант Дж. 2001; 286 (1): 1–12. [PubMed] [Академия Google]
- Гассемиан М., Намбара Э., Катлер С., Кавайде Х., Камия Ю., Маккорт П. Регуляция передачи сигналов абсцизовой кислоты посредством пути ответа на этилен у арабидопсиса. Растительная клетка. 2000;126(1):1117–1126. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Гибсон С.И., Лаби Р.Дж., Ким Д. Нечувствительный к сахару1 (sis1) мутант арабидопсиса в аллеле ctr1. Биохим. Биофиз. Рез. Комм. 2001; 2806(1):196–203. [PubMed] [Google Scholar]
- Giraudat J., Hauge B.M., Valon C., Smalle J., Parcy F., Goodman H.M. Изоляция гена ABI3 арабидопсиса путем позиционного клонирования. Растительная клетка. 1992;46(1):1251–1261. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Гирке Т., Тодд Дж., Рууска С., Уайт Дж., Беннинг С., Олрогге Дж. Микрочиповый анализ развивающихся семян арабидопсиса. Завод Физиол. 2000;1246(1):1570–1581. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Гольдберг Р. Б. , Пайва Г., Ядегари Р. Эмбриогенез растений: от зиготы к семени. Наука. 1994;2666(1):605–614. [PubMed] [Google Scholar]
- Грэм И. А. Мобилизация масла для хранения семян. Анну. Преподобный завод биол. 2008;596(1):115–142. [PubMed] [Google Scholar]
- Griffiths J., Murase K., Rieu I., Zentella R., Zhang Z.L., Powers S.J., Gong F., Phillips A.L., Hedden P., Sun T.P., Thomas S.G. Генетическая характеристика и функциональный анализ рецепторов гиббереллина GID1 у арабидопсиса. Растительная клетка. 2006;186(1):3399–3414. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Gualberti G., Papi M., Bellucci L., Ricci I., Bouchez D., Camilleri C., Costantino P., Vittorioso P. Мутации в цинке Dof гены пальцев DAG2 и DAG1 влияют противоположным образом на прорастание семян арабидопсиса. Растительная клетка. 2002;146(1):1253–1263. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Guilfoyle TJ, Hagen G. Факторы отклика на ауксин. Курс. мнение биол. растений 2007;106(1):453–460. [PubMed] [Google Scholar]
- Gutierrez L., Van Wuytswinkel O., Castelain M., Bellini C. Комбинированные сети, регулирующие созревание семян. Тенденции Растениевод. 2007;126(1):294–300. [PubMed] [Академия Google]
- Hattori T., Vasil V., Rosenkrans L., Hannah L.C., McCarty D.R., Vasil I.K. Ген Viviparous-1 и абсцизовая кислота активируют регуляторный ген C1 для биосинтеза антоцианов во время созревания семян кукурузы. Гены Дев. 1992;66(1):609–618. [PubMed] [Google Scholar]
- Хаяси М., Торияма К., Кондо М., Нисимура М. Устойчивые к 2,4-дихлорфеноксимасляной кислоте мутанты арабидопсиса имеют дефекты бета-окисления глиоксисомальных жирных кислот. Растительная клетка. 1998;106(1):183–195. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Хейс Р. Г., Кляйн В. Х. Влияние света на качество спектра во время развития растений Arabidopsis thaliana в регуляции прорастания семян. Физиология клеток растений. 1974;156(1):643–653. [Google Scholar]
- Хеллиуэлл С. А., Шелдон С. С., Олив М. Р., Уокер А. Р., Зееваарт Дж. А. Д., Пикок У. Дж., Деннис Э. С. Клонирование гена энткауреноксидазы арабидопсиса GA3. проц. Натл. акад. науч. США. 1998;956(1):9019–9024. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Хефер А., Робертс Дж. А. Контроль прорастания семян у Trollius ledebouri: нарушение покоя. Планта. 1985;1666(1):314–320. [PubMed] [Google Scholar]
- Hilhorst HW Определения и гипотезы покоя семян. В: Брэдфорд К., Ноногаки Х., редакторы. Развитие семян, покой и прорастание. 1. Том. 6. Оксфорд, Великобритания: издательство Blackwell Publishing; 2007. С. 50–67. [Google Scholar]
- Хирано К., Уэгути-Танака М., Мацуока М. GID1-опосредованная передача сигналов гиббереллина в растениях. Тенденции Растениевод. 2008;136(1):192–199. [PubMed] [Google Scholar]
- Холдсворт М. Дж., Бенцинк Л., Соппе В. Дж. Молекулярные сети, регулирующие созревание, дозревание, покой и прорастание семян арабидопсиса. Новый Фитол. 2008а; 1796(1):33–54. [PubMed] [Google Scholar]
- Холдсворт М. Дж., Финч-Сэвидж В. Э., Грэппин П., Джоб Д. Постгеномное исследование покоя семян и прорастания. Тенденции Растениевод. 2008b;136(1):7–13. [PubMed] [Google Scholar]
- Hong S. H., Vierling E. Hsp101 необходим для устойчивости к жаре, но необязателен для развития и прорастания в отсутствие стресса. Плант Дж. 2001; 266 (1): 1–12. [PubMed] [Google Scholar]
- Hugouvieux V., Kwak JM, Schroeder JI. Белок, связывающий кэп мРНК, ABh2, модулирует раннюю передачу сигнала абсцизовой кислоты у Arabidopsis. Клетка. 2001;1066(1):477–487. [PubMed] [Академия Google]
- Huijser C., Kortstee A.J., Pego JV., Weisbeek P., Wisman E., Smeekens S. Ген SUCROSE UNCOUPLED-6 арабидопсиса идентичен гену нечувствительного к абсцизовой кислоте-4: участие абсцизовой кислоты в реакциях на сахар. Плант Дж. 2000; 236 (1): 577–585. [PubMed] [Google Scholar]
- Юти С., Судзуки Х., Ким Ю. К., Юти А., Куромори Т., Уэгучи-Танака М., Асами Т., Ямагути И. , Мацуока М., Кобаяши М., Накадзима М. Множественная потеря функции рецептора гиббереллина арабидопсиса AtGID1s полностью отключает сигнал гиббереллина. Плант Дж. 2007; 506(1):958–966. [PubMed] [Google Scholar]
- Jacobsen S.E., Binkowski K.A., Olszewski N.E. SPINDLY, белок тетратрикопептидных повторов, участвующий в передаче сигнала гиббереллина у арабидопсиса. проц. Натл. акад. науч. США. 1996;936(1):9292–9296. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Johnson C.S., Kolevski B., Smyth D.R. TRANSPARENT TESTA GLABRA2, ген развития трихом и семенной кожуры арабидопсиса, кодирует фактор транскрипции Wrky. Растительная клетка. 2002;146(1):1359–1375. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Кагая Ю., Тойосима Р., Окуда Р., Усуи Х., Ямамото А., Хаттори Т. ЛИСТОВАЯ СЕМЯДЕЛЬКА1 контролирует гены запасных белков семян посредством регуляции FUSCA3 и АБСЦИЗОНОВОЙ КИСЛОТЫ INSEN-SITIVE3. Физиология клеток растений. 2005;466(1):399–406. [PubMed] [Google Scholar]
- Karssen C. M. Поглощение и действие абсцизовой кислоты во время индукции и прогрессирования роста корешков в семенах Chenopodium album. Физиол. Завод. 1976;366(1):259–263. [Google Scholar]
- Karssen C.M., Brinkhorst-van der Swan D.L.C., Breekland A.E., Koornneef M. Индукция покоя во время развития семян эндогенной абсцизовой кислотой: исследования генотипов Arabidopsis thaliana (L.) с дефицитом абсцизовой кислоты Heynh. Планта. 1983;1576(1):158–165. [PubMed] [Google Scholar]
- Karssen C.M., Zagórski S., Kepczynski J., Groot S.P.C. Ключевая роль эндогенных гиббереллинов в контроле прорастания семян. Анна. Бот. 1989;636(1):71–80. [Google Scholar]
- Катагири Т., Исияма К., Като Т., Табата С., Кобаяси М., Шинозаки К. Важная роль фосфатидной кислоты в передаче сигналов АБК во время прорастания у Arabidopsis thaliana. Плант Дж. 2005; 436 (1): 107–117. [PubMed] [Google Scholar]
- Кейт К., Крамл М., Денглер Н. Г., Маккорт П. Фуска3: Гетерохронная мутация, влияющая на позднее развитие эмбриона у арабидопсиса. Растительная клетка. 1994;66(1):589–600. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Kepczynski J., Kepczynska E. Этилен в состоянии покоя и прорастания семян. Физиол. Завод. 1997;1016(1):720–726. [Google Scholar]
- Кибер Дж. Дж., Ротенберг М., Роман Г., Фельдманн К. А., Экер Дж. Р. CTR1, негативный регулятор пути ответа на этилен у арабидопсиса, кодирует представителя семейства протеинкиназ raf. Клетка. 1993;726(1):427–441. [PubMed] [Google Scholar]
- Kim D. H., Yamaguchi S., Lim S., Oh E., Park J., Hanada A., Kamiya Y., Choi G. SOMNUS, белок цинковых пальцев CCCH-типа у арабидопсиса , отрицательно регулирует светозависимое прорастание семян после PIL5. Растительная клетка. 2008;206(1):1260–1277. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Китамура С., Шиказоно Н., Танака А. Субклеточная локализация TT19, который участвует в накоплении флавоноидов у арабидопсиса. Физиология клеток растений. 2004;456(1):S180. [Google Scholar]
- Ко Дж. Х., Ян С. Х., Хан К. Х. Повышенная регуляция гена RING-h3 арабидопсиса, XERICO, придает засухоустойчивость за счет увеличения биосинтеза абсцизовой кислоты. Плант Дж. 2006; 476 (1): 343–355. [PubMed] [Google Scholar]
- Кохчи Т., Мукоугава К., Франкенберг Н., Масуда М., Йокота А., Лагариас Дж. К. Ген арабидопсиса HY2 кодирует фитохромомобилинсинтазу, ферредоксин-зависимую биливердинредуктазу. Растительная клетка. 2001;136(1):425–436. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Koornneef M., Van der Veen J.H. Индукция и анализ чувствительных к гиббереллину мутантов в Arabidopsis thaliana (L.) Heynh. Теор. заявл. Жене. 1980;586(1):257–263. [PubMed] [Google Scholar]
- Koornneef M., Reuling G., Karssen C.M. Выделение и характеристика нечувствительных к абсцизовой кислоте мутантов Arabidopsis thaliana. Физиол. Завод. 1984;616(1):377–383. [Google Scholar]
- Koornneef M., Hanhart C.J., Hilhorst H.W.M., Karssen C.M. In vivo ингибирование развития семян и накопление резервного белка в рекомбинантах биосинтеза абсцизовой кислоты и мутантов реактивности у Arabidopsis thaliana. Завод Физиол. 1989;906(1):463–469. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Koornneef M., Karssen C.M. Покой семян и прорастание. В: Meyerowitz EM, Somerville CR, редакторы. Арабидопсис. 1. Том. 6. Колд-Спринг-Харбор: Лабораторный пресс Колд-Спринг-Харбор; 1994. С. 313–334. [Google Scholar]
- Корннеф М., Алонсо-Бланко К., Бенцинк Л., Бланкенстин-де Врис Х., Дебожон И., Ханхарт С. Дж., Леон-Клоостерзил К. М., Петерс Т., Раз В. Генетика семян покоя у Arabidopsis thaliana. В: Viémont JD, Crabbé J., editors. Покой у растения. 1. Том. 6. КАБ Интернэшнл; Уоллингфорд, Великобритания: 2000. стр. 365–373. [Академия Google]
- Котак С., Вирлинг Э., Баумляйн Х., фон Коскулл-Доринг П. Новый транскрипционный каскад, регулирующий экспрессию белков теплового стресса во время развития семян арабидопсиса. Растительная клетка. 2007;196(1):182–195. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Крок Б., Шмидт С., Хертвек С., Болдуин И. Т. Абсцизовая кислота растительного происхождения и четыре терпена обеспечивают покой семян послепожарного однолетника Nicotiana attenuata. Семенная наука. Рез. 2002;126(1):239–252. [Академия Google]
- Крой Т., Савино Г., Валон С., Жиродат Дж., Парси Ф. Регуляция экспрессии генов запасных белков у арабидопсиса. Разработка. 2003;1306(1):6065–6073. [PubMed] [Google Scholar]
- Кучера Б., Кон М. А., Лейбнер-Метцгер Г. Взаимодействие гормонов растений во время выхода семян из состояния покоя и прорастания. Семенная наука. Рез. 2006;156(1):281–307. [Google Scholar]
- Куглер И. Untersuchungen über das Keimverhalten einiger Rassen von Arabidopsis thaliana (L.) Heynh. Ein Beitrag zum Problem der Lichtkeimung. Бейтр. биол. Пфланцен. 1951;286(1):211–243. [Google Scholar]
- Кусиро Т., Окамото М., Накабаяси К., Ямагиши К., Китамура С., Асами Т., Хираи Н., Косиба Т., Камия Ю., Намбара Э. Цитохром P450 CYP707A арабидопсиса кодирует 8′-гидроксилазы АБК: ключевые ферменты катаболизма АБК. EMBO J. 2004; 236 (1): 1647–1656. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Kwong R. W., Bui A. Q. , Lee H., Kwong L. W., Fischer R. L., Goldberg R. B., Harada J. J. LEAFY COTYLEDON1-LIKE определяет класс регуляторов, необходимых для развития эмбриона. Растительная клетка. 2003;156(1):5–18. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Лаби Р. Дж., Кинкейд С., Ким Д., Гибсон С. И. Нечувствительные к сахару мутанты арабидопсиса sis4 и sis5 дефектны в синтезе абсцизовой кислоты и ответной реакции. Плант Дж. 2000; 236 (1): 587–596. [PubMed] [Google Scholar]
- Ламаттина Л., Гарсия-Мата К., Грациано М., Паннуссат Г. Оксид азота: универсальность обширной сигнальной молекулы. Анну. Преподобный завод биол. 2003; 546(1):109–136. [PubMed] [Google Scholar]
- Lee S., Cheng H., King K.E., Wang W., He Y., Hussain A., Lo J., Harberd NP, Peng J. Гиббереллин регулирует прорастание семян арабидопсиса через RGL2, GAI/RGA-подобный ген, экспрессия которого повышается после впитывания. Гены Дев. 2002;166(1):646–658. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Лефевр В. , Норт Х., Фрей А., Сотта Б., Сео М., Окамото М., Нам-бара Э., Марион-Полл А. Функциональный анализ генов NCED6 и NCED9 арабидопсиса показывает, что АБК синтезируется в эндосперм участвует в индукции покоя семян. Плант Дж. 2006; 456 (1): 309–319. [PubMed] [Google Scholar]
- Леон П., Шин Дж. Связь сахара и гормонов. Тенденции Растениевод. 2003;86(1):110–116. [PubMed] [Google Scholar]
- Лепинец Л., Дебожон И., Рутабул Дж. М., Бодри А., Пурсель Л., Неси Н., Кабош М. Генетика и биохимия флавоноидов семян. Анну. Преподобный завод биол. 2006;576(1):405–430. [PubMed] [Академия Google]
- Лейбнер-Мецгер Г., Фрюндт К., Фёгели-Ланге Р., Майнс Ф. b-1,3-глюканазы класса I в эндосперме табака при прорастании. Завод Физиол. 1995;1096(1):751–759. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Лейбнер-Мецгер Г. Функции и регуляция бета-1,3-глюканаз во время прорастания семян, выхода из покоя и после созревания. Семенная наука. Рез. 2003;136(1):17–34. [Google Scholar]
- Leubner-Metzger G. Экспрессия гена бета-1,3-глюканазы в малогидратированных семенах как механизм выхода из состояния покоя во время дозревания табака. Плант Дж. 2005; 416 (1): 133–145. [PubMed] [Академия Google]
- Leung J., Bouvier-Durand M., Morris P.C., Guerrier D., Chefdor F., Giraudat J. Ген ABA-ответа арабидопсиса ABI1: особенности кальций-модулируемой протеинфосфатазы. Наука. 1994;2646(1):14481–1452. [PubMed] [Google Scholar]
- Leung J., Merlot S., Giraudat J. Ген ABI2 арабидопсиса является гомологом ABI1 и участвует в передаче сигнала абсцизовой кислоты избыточной протеиновой серин/треонинфосфатазы 2C. Растительная клетка. 1997; 96(1):759–771. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Леунг Дж., Жиродат Дж. Клонирование генов Arabidopsis thaliana путем перемещения хромосом. Методы Мол. биол. 1998;826(1):277–303. [PubMed] [Google Scholar]
- Леон-Клоостерзил К. М., Кейзер С. Дж., Коорннеф М. Мутант арабидопсиса в форме семени, пораженный покровным покровом. Растительная клетка. 1994;66(1):385–392. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Леон-Клоостерзил К. М., ван де Бант Г. А., Зееваарт Дж. А. Д., Коорннеф М. Мутанты арабидопсиса с уменьшенным периодом покоя семян. Завод Физиол. 1996а; 1106(1):233–240. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Леон-Клоостерзиэль К. М., Гил М. А., Руйс Г. Дж., Якобсен С. Э., Ольшевски Н. Э., Шварц С. Х., Зееваарт Дж. А. Д., Коорннеф М. Выделение и характеристика абсцизовой кислоты-дефицитных мутантов арабидопсиса в двух новых местах. Плант Дж. 1996b; 106 (1): 655–661. [PubMed] [Google Scholar]
- Ли Дж., Нагпал П., Витарт В., МакМоррис Т. С., Чори Дж. Роль брассиностероидов в светозависимом развитии арабидопсиса. Наука. 1996;2726(1):398–401. [PubMed] [Google Scholar]
- Ли Дж., Чори Дж. Предполагаемая киназа рецептора повторов, богатых лейцином, участвует в передаче сигнала брассиностероидов. Клетка. 1997;906(1):825–827. [PubMed] [Google Scholar]
- Лю П. П., Коидзука Н., Хомриххаузен Т М., Хьюитт Дж. Р., Мартин Р. К., Ноногаки Х. Крупномасштабный скрининг линий энхансеров-ловушек арабидопсиса на гены, связанные с прорастанием семян. Плант Дж. 2005; 416 (1): 936–944. [PubMed] [Google Scholar]
- Лю П. П., Монтгомери Т. А., Фальгрен Н., Касшау К. Д., Ноногаки Х., Каррингтон Дж. К. Подавление AUXIN RESPONSE FACTOR10 с помощью микроРНК160 имеет решающее значение для прорастания семян и после прорастания. Плант Дж. 2007; 526 (1): 133–146. [PubMed] [Академия Google]
- Liu Y., Koornneef M., Soppe W. J. Отсутствие моноубиквитинирования гистона h3B в мутанте Arabidopsis hub1 (rdo4) указывает на роль ремоделирования хроматина в состоянии покоя семян. Растительная клетка. 2007;196(1):433–444. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Лопес-Молина Л., Монгранд С., Чуа Н. Х. Контрольная точка задержки развития после прорастания опосредуется абсцизовой кислотой и требует фактора транскрипции ABI5 у арабидопсиса. проц. Натл. акад. науч. США. 2001;986(1):4782–4787. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Lotan T., Ohto M., Yee K.M., West M.A., Lo R., Kwong R.W., Yamagishi K., Fischer R.L., Goldberg R.B., Harada J.J. Arabidopsis LEAFY COTYLEDON1 достаточно для индукции развития эмбриона в вегетативных клетках. Клетка. 1998;936(1):1195–1205. [PubMed] [Google Scholar]
- Luerssen H., Kirik V., Herrmann P., Misera S. FUSCA3 кодирует белок с консервативным VP1/AB13-подобным доменом B3, который имеет функциональное значение для регуляции созревания семян в Талианский арабидопсис. Завод Ж. 1998;156(1):755–764. [PubMed] [Google Scholar]
- Macquet A., Ralet M.C., Loudet O., Kronenberger J., Mouille G., Marion-Poll A., North H.M. Естественная мутация в образце Arabidopsis влияет на бета-D- галактозидаза, которая увеличивает гидрофильный потенциал рамногалактуронана I в слизи семян. Растительная клетка. 2007;196(1):3990–4006. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Манц Б. , Мюллер К., Кучера Б., Фольке Ф., Лейбнер-Мецгер Г. Поглощение и распределение воды в прорастающих семенах табака исследовано in vivo с помощью ядерного магнитного резонанса визуализация. Завод Физиол. 2005;1386(1):1538–1551. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Masubelele N. H., Dewitte W., Menges M., Maughan S., Collins C., Huntley R., Nieuwland J., Scofield S., Murray J. A. H. Циклины D-типа активируют деление в верхушке корня, способствуя прорастанию семян в Арабидопсис. проц. Натл. акад. науч. США. 2005;1026(1):15694–15699. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Mayer U., Ruiz RAT, Berleth T., Misera S., Jürgens G. Мутации, влияющие на организацию тела у эмбриона арабидопсиса. Природа. 1991;3536(1):402–407. [Академия Google]
- Маццелла М. А., Арана М. В., Станелони Р. Дж., Перельман С., Родригес Батиллер М. Дж., Мускетти Дж., Сердан П. Д., Чен К., Санчес Р. А., Чжу Т., Чори Дж., Казал Дж. Дж. Фитохромный контроль транскриптома арабидопсиса предвосхищает воздействие света на рассаду. Растительная клетка. 2005;176(1):2507–2516. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Макаби Дж. М., Хилл Т. А., Скиннер Д. Дж., Ижаки А., Хаузер Б. А., Мейстер Р. Дж., Редди Г. В., Мейеровиц Э. М., Боуман Дж. Л., Гассер К. С. АБЕРРАНТНАЯ ТЕСТА ФОРМА кодирует канадский член семьи, связывая определение полярности с разделением и ростом покровов семязачатка арабидопсиса. Плант Дж. 2006; 466 (1): 522–531. [PubMed] [Академия Google]
- McCullough J.M., Shropshire W.J. Физиологическая предопределение реакции прорастания у Arabidopsis thaliana (L.) Heynh. Физиология клеток растений. 1970;116(1):139–148. [Google Scholar]
- McGinnis K.M., Thomas S.G., Soule JD., Strader L.C., Zale J.M., Sun T.P., Steber C.M. Ген арабидопсиса SLEEPY1 кодирует предполагаемую субъединицу F-box убиквитинлигазы SCF E3. Растительная клетка. 2003;156(1):1120–1130. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Meinke D. W. Развитие семян в Arabidopsis thaliana. В: Meyerowitz EM, Somerville CR, редакторы. Арабидопсис. 1. Том. 6. Колд-Спринг-Харбор: Лабораторный пресс Колд-Спринг-Харбор; 1994. стр. 253–298. [Google Scholar]
- Мейнке Д. В. Молекулярная генетика эмбриогенеза растений. Анну. Преподобный Завод Физиол. Завод Мол. биол. 1995;466(1):369–394. [Google Scholar]
- Мэн П.-Х., Маке А., Лудет О., Марион-Полл А., Норт Х. Анализ естественной аллельной вариации, контролирующей всхожесть семян Arabidopsis thaliana в ответ на холод и темноту: выявление трех основных количественных признаков локусы Мол. Завод. 2008;16(1):145–154. [PubMed] [Google Scholar]
- Meyer K., Leube M.P., Grill E. Протеинфосфатаза 2C, участвующая в передаче сигнала ABA у Arabidopsis thaliana. Наука. 1994;2646(1):1452–1455. [PubMed] [Google Scholar]
- Митчем М. Г., Ямагути С., Ханада А., Кувахара А., Йошиока Ю., Като Т., Табата С., Камия Ю., Сун Т. П. Различные и перекрывающиеся роли двух гиббереллинов 3 -оксидазы в развитии арабидопсиса. Плант Дж. 2006; 456 (1): 804–818. [PubMed] [Google Scholar]
- Мюллер А. , Гуан С., Галвейлер Л., Танцлер П., Хейсер П., Маршан А., Парри Г., Беннетт М., Висман Э., Пальме К. AtPIN2 определяет локус арабидопсиса для контроля корневого гравитропизма. EMBO J. 1998;176(1):6903–6911. [Статья бесплатно PMC] [PubMed] [Google Scholar]
- Muller K., Tintelnot S., Leubner-Metzger G. Ограниченное эндоспермом прорастание семян Brassicaceae: абсцизовая кислота ингибирует индуцированное эмбрионом ослабление эндосперма Lepidium sativum (кресс-салат) и эндосперма разрыв кресс-салата и Arabidopsis thaliana. Физиология клеток растений. 2006;476(1):864–877. [PubMed] [Google Scholar]
- Мунир Дж., Дорн Л. А., Донохью К., Шмитт Дж. Влияние материнского фотопериода на сезонный покой у Arabidopsis thaliana (Brassicaceae). амер. жур. Бот. 2001;886(1):1240–1249. [PubMed] [Google Scholar]
- Muramoto T., Kohchi T., Yokota A., Hwang I., Goodman H.M. Фотоморфогенный мутант hy1 арабидопсиса дефицитен в биосинтезе хромофора фтиохрома в результате мутации в пластиде. гемоксигеназы. Растительная клетка. 1999;116(1):335–347. [Статья бесплатно PMC] [PubMed] [Google Scholar]
- Накабаяши К., Окамото М., Косиба Т., Камия Ю., Намбара Э. Полногеномное профилирование хранимой мРНК в прорастании семян Arabidopsis thaliana: эпигенетическая и генетическая регуляция транскрипции в семени. Плант Дж. 2005; 416 (1): 69.7–709. [PubMed] [Google Scholar]
- Nambara E., Naito S., McCourt P. Мутант арабидопсиса, дефектный в развитии семян и накоплении запасного белка, представляет собой новый аллель abi3. Плант Дж. 1992; 26 (1): 435–441. [Google Scholar]
- Nambara E., Hayama R., Tsuchiya Y., Nishimura M., Kawaide H., Kamiya Y., Naito S. Роль локусов ABI3 и FUS3 в Arabidopsis thaliana в фазовом переходе от позднего эмбрионального развития к прорастанию. Девел. биол. 2000;2206(1):412–423. [PubMed] [Академия Google]
- Nesi N., Debaujon I., Jond C., Pelletier G., Caboche M., Lepiniec L. Ген TT8 кодирует основной белок домена спираль-петля-спираль, необходимый для экспрессии генов DFR и BAN у стручков арабидопсиса. Растительная клетка. 2000; 126 (1): 1863–1878. [Статья бесплатно PMC] [PubMed] [Google Scholar]
- Nesi N., Jond C., Debaujon I., Caboche M., Lepiniec L. Ген TT2 арабидопсиса кодирует белок домена MYB R2R3, который действует как ключевой детерминант накопление в развивающихся семенах. Растительная клетка. 2001;136(1):2099–2114. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Nesi N., Debaujon I., Jond C., Stewart A.J., Jenkins G.I., Caboche M., Lepiniec L. Локус TRANSPARENT TESTA16 кодирует домен MADS ARABIDOPSIS B SISTER белка и необходим для правильного развития и пигментации семенной кожуры. Растительная клетка. 2002;146(1):2463–2479. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Ни Д. А., Ван Л. Дж., Дин С. Х., Сюй З. Х. Распределение и транспорт ауксина во время эмбриогенеза и прорастания семян арабидопсиса. Сотовый рез. 2001;116(1):273–278. [PubMed] [Академия Google]
- Нисимура Н., Йошида Т., Мураяма М., Асами Т., Шинозаки К., Хираяма Т. Выделение и характеристика новых мутантов, влияющих на чувствительность к абсцизовой кислоте прорастания и роста проростков арабидопсиса. Физиология клеток растений. 2004;456(1):1485–1499. [PubMed] [Google Scholar]
- Нисимура Н., Йошида Т., Китахата Н., Асами Т., Шинозаки К., Хираяма Т. ABA-Hypersensive Germination1 кодирует протеинфосфатазу 2C, важный компонент передачи сигналов абсцизовой кислоты в Семена арабидопсиса. Плант Дж. 2007; 506(1):935–949. [PubMed] [Google Scholar]
- Niu X. P., Helentjaris T., Bate N. J. Кукуруза AB14 связывает связующий элемент 1 в генах ответа на абсцизовую кислоту и сахар. Растительная клетка. 2002;146(1):2565–2575. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- North H.M., De Almeida A., Boutin JP, Frey A., To A., Botran L., Sotta B., Marion-Poll A. The Arabidopsis ABA- Дефицитный мутант aba4 демонстрирует, что основной путь индуцированного стрессом накопления АБК проходит через изомеры неоксантина. Плант Дж. 2007; 506 (1): 810–824. [PubMed] [Академия Google]
- Огава М., Ханада А., Ямаути Ю., Кувахара А., Камия Ю., Я-магучи С. Биосинтез гиббереллина и реакция во время прорастания семян арабидопсиса. Растительная клетка. 2003;156(1):1591–1604. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Oh E., Kim J., Park E., Kim JI, Kang C., Choi G. PIL5, взаимодействующий с фитохромом основной белок спираль-петля-спираль, является ключевым негативным регулятором прорастания семян у Arabidopsis thaliana. Растительная клетка. 2004;166(1):3045–3058. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Oh E., Yamaguchi S., Kamiya Y., Bae G., Chung W.I., Choi G. Свет активирует деградацию белка PIL5, чтобы способствовать прорастанию семян через гиббереллин в арабидопсисе. Плант Дж. 2006; 476 (1): 124–139. [PubMed] [Google Scholar]
- О Э., Ямагути С., Ху Дж., Юсуке Дж., Юнг Б., Пайк И., Ли Х. С., Сун Т. П., Камия Ю., Чой Г. PIL5, фитохром — взаимодействующий белок bHLH, регулирует чувствительность к гиббереллину, связываясь непосредственно с промоторами GAI и RGA в семенах арабидопсиса. Растительная клетка. 2007;196(1):1192–1208. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Окамото М., Кувахара А., Сео М., Куширо Т., Асами Т., Хираи Н., Камия Ю., Косиба Т., Намбара Э. CYP707A1 и CYP707A2, которые кодируют 8′-гидроксилазы абсцизовой кислоты, необходимы для надлежащего контроля покоя семян и прорастания у арабидопсиса. Завод Физиол. 2006;1416(1):97–107. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Ooms JJJ, Léon-Kloosterziel KM, Bartels D., Koornneef M., Karssen CM Приобретение устойчивости к высыханию и долговечности семян Arabidopsis thaliana — сравнительное исследование с использованием абсцизовой кислоты — бесчувственные мутанты abi3. Завод Физиол. 1993;1026(1):1185–1191. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Пандей С., Чен Дж. Г., Джонс А. М., Ассманн С. М. Мутанты комплекса G-белка гиперчувствительны к абсцизовой кислоте, регулирующей прорастание и развитие после прорастания. Завод Физиол. 2006;1416(1):243–256. [Статья бесплатно PMC] [PubMed] [Google Scholar]Retracted
- Papi M., Sabatini S., Bouchez D., Camilleri C., Costantino P., Vittorioso P. Идентификация и нарушение гена цинкового пальца арабидопсиса, контролирующего прорастание семян . Гены Дев. 2000;146(1):28–33. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Папонов И. А., Тил В. Д., Требар М., Блилоу К., Пальме К. Ускорители оттока ауксина PIN: эволюционные и функциональные перспективы. Тенденции пл. науч. 2005;106(1):170–177. [PubMed] [Google Scholar]
- Parcy F., Valon C., Raynal M., Gaubier-Comella P., Delseny M., Giraudat J. Регуляция программ экспрессии генов во время развития семян арабидопсиса: роли локуса ABI3 и эндогенной абсцизовой кислоты. Растительная клетка. 1994; 66 (1): 1567–1582. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Парси Ф., Жиродат Дж. Взаимодействия между ABI1 и эктопически экспрессируемыми генами ABI3 в контроле реакции абсцизовой кислоты в вегетативных тканях арабидопсиса. Плант Дж. 1997; 116 (1): 693–702. [PubMed] [Google Scholar]
- Петерс А. Дж., Бланкестин-Де Врис Х., Ханхарт С. Дж., Леон-Клоостерзил К. М., Зееваарт Дж. А., Корннеф М. Характеристика мутантов с уменьшенным периодом покоя семян в двух новых локусах rdo и дальнейшая характеристика rdo1 и rdo2 у арабидопсиса. Физиол. Завод. 2002;1156(1):604–612. [PubMed] [Академия Google]
- Пеллетье М.К., Ширли Б.В. Анализ флаванон-3-гидроксилазы в проростках арабидопсиса. Координация регуляции с помощью халконсинтазы и халконизомеразы. Завод Физиол. 1996;1116(1):339–345. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Penfield S., Rylott E.L., Gilday A.D., Graham S., Larson T.R., Graham I.A. Резервная мобилизация эндосперма арабидопсиса способствует удлинению гипокотиля в темноте, не зависит от абсцизной кислоты и требует фосфоенолпируваткарбоксикиназы1. Растительная клетка. 2004;166(1):2705–2718. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Penfield S., Josse E. M., Kannangara R., Gilday A.D., Halliday KJ, Graham I.A. Прорастание семян контролируется холодом и светом с помощью транскрипционного фактора bHLH SPATULA. Курс. биол. 2005;156(1):1998–2006. [PubMed] [Google Scholar]
- Penfield S., Li Y., Gilday A.D., Graham S., Graham I.A. Arabidopsis ABA INSENSITIVE4 регулирует мобилизацию липидов в зародыше и выявляет подавление прорастания семян эндоспермом. Растительная клетка. 2006; 186 (1): 1887–1899. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Брэдфорд К., Ноногаки Х., редакторы. Липидный обмен в период покоя семян. 1. Том. 6. Оксфорд, Великобритания: издательство Blackwell Publishing; 2007. стр. 131–152. [Google Scholar]
- Петруцелли Л., Мюллер К., Герман К., Лейбнер-Мецгер Г. Отличительные особенности экспрессии бета-1,3-глюканаз и хитиназ при прорастании семян пасленовых. Семенная наука. Рез. 2003;136(1):139–153. [Google Scholar]
- Poppe C., Schäfer E. Прорастание семян двойных мутантов Arabidopsis thaliana phyA/phyB находится под контролем фитохрома. Завод Физиол. 1997;1146(1):1487–1492. [Статья бесплатно PMC] [PubMed] [Google Scholar]
- Pourcel L., Routaboul J.M., Kerhoas L., Caboche M., Lepiniec L., Debaujon I. TRANSPARENT TESTA10 кодирует лакказоподобный фермент, участвующий в окислительной полимеризации. флавоноидов в кожуре семян арабидопсиса. Растительная клетка. 2005;176(1):2966–2980. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Pracharoenwattana I., Cornah JE, Smith S.M. Пероксисомальная цитратсинтаза арабидопсиса необходима для дыхания жирных кислот и прорастания семян. Растительная клетка. 2005;176(1):2037–2048. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Rajjou L., Gallardo K., Debaujon I., Vandekerckhove J., Job C., Job D. Влияние альфа-аманитина на протеом семян арабидопсиса выдвигает на первый план различные роли сохраненных и вновь синтезированных мРНК во время прорастания. Завод Физиол. 2004;1346(1):1598–1613. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Ratcliffe D. Характеристики прорастания и их межпопуляционная и внутрипопуляционная изменчивость у арабидопсиса. араб. Инф. Серв. 1976;136(1):34–45. [Google Scholar]
- Раз В., Бергервоет Дж. Х., Корнниф М. Последовательные шаги для остановки развития семян арабидопсиса. Разработка. 2001;1286(1):243–252. [PubMed] [Академия Google]
- Rock C.D., Zeevaart J.A.D. У мутанта aba Arabidopsis thaliana нарушен биосинтез эпоксикаротиноидов. проц. Натл. акад. науч. США. 1991;886(1):7496–7499. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Rock CD Пути к экспрессии генов, регулируемой абсцизовой кислотой. Новый Фитол. 2000;1486(1):357–396. [Google Scholar]
- Rognoni S., Teng S., Arru L., Smeekens S.C.M., Perata P. Влияние сахара на раннее развитие проростков арабидопсиса. Регламент роста растений 2007;526(1):217–228. [Академия Google]
- Rook F., Corke F., Card R., Munz G., Smith C., Bevan M.W. Мутанты с нарушением индукции сахарозы обнаруживают модуляцию индуцированной сахаром экспрессии генов биосинтеза крахмала посредством передачи сигналов абсцизовой кислоты. Плант Дж. 2001; 266 (1): 421–433. [PubMed] [Google Scholar]
- Рук Ф., Хадингем С.А., Ли Ю., Беван М.В. Пути реакции на сахар и ABA и контроль экспрессии генов. Окружающая среда растительных клеток. 2006;296(1):426–434. [PubMed] [Google Scholar]
- Рассел Л., Ларнер В., Куруп С., Бугурд С., Холдсворт М. Дж. Локус COMATOSE арабидопсиса регулирует потенциал прорастания. Разработка. 2000;1276(1):3759–3767. [PubMed] [Google Scholar]
- Rylott E.L., Rogers C.A., Gilday A.D., Edgell T., Larson T.R., Graham I.A. Мутанты Arabidopsis по активности ацил-КоА-оксидазы с короткой и средней цепью накапливают ацил-КоА и обнаруживают, что жирная кислота бета-окисление необходимо для развития эмбриона. Дж. Биол. хим. 2003;2786(1):21370–21377. [PubMed] [Google Scholar]
- Rylott E. L., Eastmond P. J., Gilday A. D., Slocombe S. P., Larson T. R., Baker A., Graham I. A. Ген пероксисомального бета-окисления (MFP2) Arabidopsis thaliana MULTI-FUNCTIONAL PROTEIN (MFP2) необходим для рассады учреждение. Плант Дж. 2006; 456(1):930–941. [PubMed] [Google Scholar]
- Саез А., Роберт Н., Мактаби М. Х., Шредер Дж. И., Серрано Р., Родригес П. Л. Повышение чувствительности к абсцизовой кислоте и снижение потребления воды у арабидопсиса путем комбинированной инактивации протеинфосфатаз типа 2С ABI1 и HAB1. Завод Физиол. 2006;1416(1):1389–1399. [Статья бесплатно PMC] [PubMed] [Google Scholar]
- Sagasser M., Lu G.H., Hahlbrock K., Weisshaar B.A. thaliana TRANSPARENT TESTA 1 участвует в развитии семенной кожуры и определяет подсемейство WIP белков цинковых пальцев растений. Джинс Девель. 2002; 166(1):138–149.. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Салаита Л., Кар Р. К., Маджи М., Дауни А. Б. Идентификация и характеристика мутантов, способных к быстрому прорастанию семян при 10 градусах Цельсия из меченых активацией линий Arabidopsis thaliana. Дж. Эксп. Бот. 2005;566(1):2059–2069. [PubMed] [Google Scholar]
- Schneitz K. Молекулярно-генетический контроль развития яйцеклетки. Курс. мнение биол. растений 1999;26(1):13–17. [PubMed] [Google Scholar]
- Schoenbohm C., Martens S., Eder C., Forkmann G., Weisshaar B. Идентификация гена флавоноид-3′-гидроксилазы Arabidopsis thaliana и функциональная экспрессия кодируемого фермента P450. биол. хим. 2000;3816(1):749–753. [PubMed] [Google Scholar]
- Schopfer P., Plachy C. Контроль прорастания семян с помощью абсцизовой кислоты. II. Влияние на поглощение воды зародышами Brassica napus L. Plant Physiol. 1984;766(1):155–160. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Schwartz SH, Léon-Kloosterziel KM, Koornneef M., Zeevaart JAD Биохимическая характеристика мутантов aba2 и aba3 в Arabidopsis thaliana. Завод Физиол. 1997;1146(1):161–166. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Шаррок Р. А., Перепел П. Х. Новые последовательности фитохромов в Arabidopsis thaliana: структура, эволюция и дифференциальная экспрессия семейства регуляторных фоторецепторов растений. Джинс Девель. 1989; 36 (1): 1745–1757. [PubMed] [Google Scholar]
- Shen Q., Ho TH. Функциональное вскрытие гена, индуцируемого абсцизовой кислотой (ABA), выявило два независимых ABA-чувствительных комплекса, каждый из которых содержит G-бокс и новый цис-действующий элемент. Растительная клетка. 1995;76(1):295–307. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Шиказоно Н., Йокота Ю., Китамура С., Судзуки С., Ватанабэ Х., Тано С., Танака А. Скорость мутаций и новые мутанты tt Arabidopsis thaliana, индуцированные углеродом. Генетика. 2003;1636(1):1449–1455. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Shinomura T., Nagatani A., Chory J., Furuya M. Индукция прорастания семян у Arabidopsis thaliana регулируется главным образом фитохромом B и, во вторую очередь, фитохромом A. Растение Физиол. 1994;1046(1):363–371. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Shinomura T., Nagatani A., Hanzawa H., Kubota M., Watanabe M., Furuya M. Спектры действия фитохрома A- и B-специфической фотоиндукции прорастания семян у Arabidopsis thaliana. проц. Натл. акад. науч. США. 1996;936(1):8129–8133. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Ширли Б.В., Хенли С., Гудман Х.М. Влияние ионизирующего излучения на геном растения: анализ двух мутаций прозрачных раковин арабидопсиса. Растительная клетка. 1992;46(1):333–347. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Скривер К., Манди Дж. Экспрессия генов в ответ на абсцизовую кислоту и осмотический стресс. Растительная клетка. 1990;26(1):503–512. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Содерман Э. М., Брокард И. М., Линч Т. Дж., Финкельштейн Р. Р. Регуляция и функция гена, нечувствительного к АБК арабидопсиса, в сигнальных сетях ответа на абсцизовую кислоту семян. Завод Физиол. 2000;1246(1):1752–1765. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Steber C. Де-подавление прорастания семян сигналами GA. В: Брэдфорд К., Ноногаки Х., редакторы. Развитие семян, покой и прорастание. 1. Том. 6. Оксфорд, Великобритания: издательство Blackwell Publishing; 2007. стр. 248–260. [Академия Google]
- Steber C.M., Cooney S., McCourt P. Выделение мутанта sly1, реагирующего на ГА, в качестве супрессора ABI1-1 у Arabidopsis thaliana. Генетика. 1998;1496(1):509–521. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Steber CM, McCourt P. Роль брассиностероидов в прорастании арабидопсиса. Завод Физиол. 2001;1256(1):763–769. [Статья бесплатно PMC] [PubMed] [Google Scholar]
- Stone S.L., Kwong L.W., Yee K.M., Pelletier J., Lepiniec L., Fischer R.L., Goldberg R.B., Harada J.J. Листовая семядоля2 кодирует фактор транскрипции домена B3, который индуцирует зародыш разработка. проц. Натл. акад. науч. США. 2001;986(1):11806–11811. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Sun T., Goodman H.M., Ausubel FM. Клонирование локуса Arabidopsis GA1 путем вычитания генома. Растительная клетка. 1992;46(1):119–128. [Статья бесплатно PMC] [PubMed] [Google Scholar]
- Teng S., Rognoni S., Bentsink L., Smeekens S. Аллель Cvi GSQ5/DOG1 арабидопсиса индуцируется ABA-опосредованным сигнальным путем сахара и повышает чувствительность к сахару. путем стимуляции экспрессии ABI4. Плант Дж. 2008; 556 (1): 372–381. [PubMed] [Академия Google]
- Тидеманн Дж., Руттен Т., Монке Г., Форвигер А., Роллетчек Х., Мейснер Д., Милковски С., Петерек С., Мок Х. П., Занк Т., Баумляйн Х. Анализ сложного семенного фенотипа : новое понимание процессов развития, регулируемых FUSCA3. Дев. биол. 2008;3176(1):1–12. [PubMed] [Google Scholar]
- А., Валон С., Савино Г., Гиллемино Дж., Девик М., Жиродат Дж., Парси Ф. Созреванием семян арабидопсиса управляет сеть локальной и избыточной регуляции генов. Растительная клетка. 2006;186(1):1642–1651. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Уэгучи-Танака М., Асикари М., Накадзима М., Ито Х., Като Э., Кобаяши М., Чоу Т. Ю., Хсинг Ю. И. К., Китано Х., Ямагути И., Мацуока М. Нечувствительный к гиббереллину карлик1 кодирует растворимый рецептор гиббереллина. Природа. 2005;4376(1):693–698. [PubMed] [Google Scholar]
- Ван дер Шаар В., Алонсо-Бланко К., Леон-Клоостерзил К. М., Янсен Р. К., Ван Оойен Дж. В., Коорнниф М. QTL-анализ покоя семян арабидопсиса с использованием рекомбинантных инбредных линий и картирование MQM . Наследственность. 1997;796(1):190–200. [PubMed] [Google Scholar]
- Vicient C. M., Bies-Etheve N., Delseny M. Изменения в экспрессии генов в листовых семядолях1 (lec1) и fusca3 (fus3) мутантов Arabidopsis thaliana L. J. Exp. Бот. 2000;516(1):995–1003. [PubMed] [Google Scholar]
- Уокер А. Р., Дэвидсон П. А., Болоньези-Уинфилд А. С., Джеймс С. М., Шринивасан Н., Бланделл Т. Л., Эш Дж. Дж., Маркс М. Д., Грей Дж. К. Локус TRANSPARENT TESTA GLABRA 1, который регулирует дифференцировку трихом и биосинтез антоцианов у арабидопсиса кодирует повторный белок WD40. Растительная клетка. 1999;116(1):1337–1349. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Вебер Х., Вобус У. Сахара как сигнальные молекулы в развитии семян растений. биол. хим. 1999;3806(1):937–944. [PubMed] [Google Scholar]
- Wehmeyer N., Vierling E. Экспрессия малых белков теплового шока в семенах отвечает на дискретные сигналы развития и предполагает общую защитную роль в устойчивости к высыханию. Завод Физиол. 2000;1226(1):1099–1108. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Wisman E., Hartmann U., Sagasser M., Baumann E., Palme K., Hahlbrock K., Saedler H., Weisshaar B. Нокаут-мутанты из мутагенизированной En-1 популяции Arabidopsis thaliana генерируют фенотипы биосинтеза фенилпропаноидов. проц. Натл. акад. науч. США. 1998;956(1):12432–12437. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Се Д.Ю., Шарма С.Б., Пайва Н.Л., Феррейра Д., Диксон Р.А. Роль антоцианидинредуктазы, кодируемой BANYULS, в биосинтезе флавоноидов растений. Наука. 2003;2996(1):396–399. [PubMed] [Google Scholar]
- Сюн Л., Ишитани М., Ли Х., Чжу Дж. К. Локус LOS5/ABA3 арабидопсиса кодирует кофактор сульферазу молибдена и модулирует экспрессию генов, чувствительных к холодовому и осмотическому стрессу. Растительная клетка. 2001;136(1):2063–2083. [Статья бесплатно PMC] [PubMed] [Google Scholar]
- Yamaguchi S., Smith MW, Brown RGS, Kamiya Y., Sun T. Регулирование фитохрома и дифференциальная экспрессия генов гиббереллин-3β-гидроксилазы в прорастающих семенах арабидопсиса. Растительная клетка. 1998;106(1):2115–2126. [Статья бесплатно PMC] [PubMed] [Google Scholar]
- Yamaguchi S., Kamiya Y., Sun T.P. Отличительные клеточно-специфические модели экспрессии ранних и поздних генов биосинтеза гиббереллина во время прорастания семян арабидопсиса. Плант Дж. 2001; 286 (1): 443–453. [PubMed] [Google Scholar]
- Ямаути Ю., Огава М., Кувахара А., Ханада А., Камия Ю., Ямагучи С. Активация биосинтеза гиббереллина и ответных путей низкой температурой во время впитывания семян Arabidopsis thaliana. Растительная клетка. 2004;166(1):367–378. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Ямаути Ю., Такэда-Камия Н., Ханада А., Огава М., Кувахара А., Сео М., Камия Ю., Ямагути С. Вклад дезактивации гиббереллина с помощью AtGA2ox2 в подавление прорастания темноволосых Семена арабидопсиса талийского. Физиология клеток растений. 2007;486(1):555–561. [PubMed] [Google Scholar]
- Ян Ю. Ю., Нагатани А., Чжао Ю. Дж., Канг Б. Дж., Кендрик Р. Э., Камия Ю. Влияние гиббереллинов на прорастание семян фитохром-дефицитных мутантов Arabidopsis thaliana. Физиология клеток растений. 1995;366(1):1205–1211. [PubMed] [Google Scholar]
- Yoine M., Ohto M. A., Onai K., Mita S., Nakamura K. Мутация lba1 геликазы РНК UPF1, участвующая в нонсенс-опосредованном распаде мРНК, вызывает плейотропные фенотипические изменения и измененную передачу сигналов сахара в Арабидопсис. Плант Дж. 2006; 476 (1): 49–62. [PubMed] [Google Scholar]
- Zeng Y., Kermode A. R. Ген голосеменного растения ABI3 функционирует в мутанте арабидопсиса, нечувствительном к абсцизовой кислоте (abi3-6), с восстановлением фенотипа дикого типа и демонстрирует сильный синергетический эффект с сахаром. в подавлении постгерминативного роста. Завод Мол. биол. 2004;566(1):731–746. [PubMed] [Академия Google]
- Зентелла Р. , Чжан З. Л., Парк М., Томас С. Г., Эндо А., Мурасе К., Флит С. М., Джикумару Ю., Намбара Э., Камия Ю., Сун Т. П. Глобальный анализ прямых мишеней della в раннем гиббереллине сигнализация у арабидопсиса. Растительная клетка. 2007;196(1):3037–3057. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Чжан Ф., Чен Г., Хуан К., Орион О., Кругман Т., Фахима Т., Король А. Б., Нево Э., Гуттерман Ю. Генетическая основа состояния покоя зерновки ячменя и устойчивости проростков к высыханию на стадии прорастания. Теор. заявл. Жене. 2005;1106(1):445–453. [PubMed] [Академия Google]
- Чжоу Л., Джанг Дж. К., Джонс Т. Л., Шин Дж. Перекрестные помехи передачи сигналов глюкозы и этилена, выявленные нечувствительным к глюкозе мутантом арабидопсиса. проц. Натл. акад. науч. США. 1998;956(1):10294–10299. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Статьи из книги Arabidopsis предоставлены здесь с разрешения Американского общества биологов растений
частей семени
частей семениЧасти семени
Бринкли Портер
2 мая 2000 г.
Краткое описание урока:
Практический урок, на котором учащиеся изучают части лимской фасоли. Учащиеся определяют семенную оболочку, зародыш и семядолю.
Уровень:
Третий и четвертый классыСтандарт содержания учебного курса:
- Описать характеристики объектов
-цвет
-гибкость
-состав
-форма
— размер
— текстура
(AL Science, Grade 3, Concept 21)
Справочная информация для учителя:
Для этого урока о семенах учитель должен ознакомиться со многими понятиями, прежде чем проводить урок.
- Лимская фасоль состоит из трех частей: кожура (семенная оболочка), зародыш (крошечное растение) и семядоля (пища).
- На каждом бобе есть маленькое белое пятнышко в том месте, где семя когда-то было прикреплено к растению, из которого оно произошло.
- Семенная кожура или кожура защищает то, что находится внутри.
- Эмбрион или крошечное растение находится внутри семенной кожуры бобов. Это часть фасоли, которая вырастет в большое растение, если семя поместить в почву и полить.
- Семядоли дают пищу крошечному растению.
Понятия, рассмотренные в уроке:
«Есть три части семени.»
«Боб или семя состоит из семенной кожуры, зародыша и семядоли».
«Зародыш — это крошечное растение, защищенное семенной оболочкой».
«Зародыш вырастет в большое растение, когда семя будет посажено в почву и
поливают.»
«Семядоли — пища для зародыша».
(AL Science, класс 3, концепция 21)
Материалы и оборудование:
*
Одна сухая лимская фасоль на ученика*Одна влажная лимская фасоль на ученика
*Одно бумажное полотенце на учащегося
*Одна копия «Big Bean» на учащегося
*Один экземпляр каждого «Внутри семени» на ученика
*Доска для сухого стирания и маркер
*Один заводской журнал на учащегося
Процедура:
- Попросите учащихся сесть на пол. Объясните учащимся, что на этом уроке они будут изучать семена растений. Задайте классу следующие вопросы и запишите ответы детей на доске:
- Нарисуйте семя на доске для сухого стирания. Посмотрите на первую страницу «Внутри семени» в детском журнале о растениях. Включите линии маркировки на свой рисунок. Спросите учащихся, как называются части растений. По мере ответа записывайте названия (семенная кожура, зародыш, семядоля) частей растения в строках для надписей.
- «Достаньте свои журналы из шкафчиков и вернитесь к своим столам.» Раздайте учащимся обе копии книги «Внутри семени». Скажите им положить листы в папку с растениями и открыть первую страницу «Внутри семени». «Заполните поля для маркировки на этой странице, а затем отложите папку в сторону».
- Раздайте бумажное полотенце, одну сухую лимскую фасоль и одну влажную лимскую фасоль каждому ребенку. Скажите учащимся следовать указаниям на второй странице «Внутри семени». Дети будут описывать, измерять и рисовать семена в соответствии с инструкциями на рабочих листах. Попросите класс заполнить страницу, за исключением описания работы внизу страницы. Они завершат эту часть после коллоквиума. Дайте учащимся 15–20 минут на работу с рабочим листом, а затем изучите части семени. Призовите их разорвать семена, заглянуть внутрь, определить части и придумать вопросы для обсуждения в классе.
- Коллоквиум (Посадите учащихся на пол): Учитель вовлекает учащихся в обсуждение, используя следующие вопросы. Учитель записывает ответы учащихся на доске.
*»Что вы знаете о семенах?»
*»Из каких частей состоит семя?»
*»Как вода может изменить семя?» (Подумайте о цвете, размере, текстуре и т. д.)
Дети будут делать прогнозы о том, как вода влияет на семена.
- Что вы заметили?
- Чем эти семена отличаются друг от друга?
- Как два семени выглядят, ощущаются, пахнут и т. д.?
- Каково назначение различных частей?
Я ожидаю, что учащиеся ответят на эти вопросы, описав, что они наблюдали во время вскрытия обоих семян. По мере того как учащиеся продолжают описывать то, что они видели, учитель снова представляет части растения. Учитель скажет: «Вы слышали, как я использовал слово «эмбрион». Кто-нибудь знает, что это слово означает?» Затем учитель скажет: «Зародыш — это часть семени, из которого вырастает крошечное растение. Семядоли — это источник пищи для растений. Оболочка семени действует как кожа для семени. важные части внутри». К концу этого обсуждения учащиеся должны были сами ответить на свои вопросы (при условии, что учитель направлял их идеи в правильном направлении) о работе частей растения.
- Верните детей за парты. В это время они уберут все кусочки семян
и бумажные полотенца. После того, как все столы будут убраны, попросите учащихся позвонить по номеру
.достаньте журналы растений и разложите их на своих столах. «Откройте свои журналы
ко второму рабочему листу Inside a Seed, над которым вы работали. Я хочу, чтобы каждый из вас,
, заполнил нижнюю часть страницы, написав задания трех исходных частей в указанных строках. Просто скажите, что делает каждая часть. Когда вы закончите, положите свой журнал обратно в каморку и сядьте на свое место».0005
Оценка:
Учитель вручит каждому ученику копию «Большой фасоли». Этот лист будет служить оценкой, но после проверки учителем будет помещен в журнал растений. Учащимся предлагается заполнить каждый пропуск правильным ответом. Каждая часть начального числа имеет две заготовки, одну для маркировки и одну для должностной инструкции. Напомните учащимся написать свои имена на листах перед тем, как сдать их учителю.
Полезные интернет-ресурсы:
http://www.lessonplanspage.com/index.htmlЭтот интернет-сайт представляет собой план урока по проращиванию семян. На уроке разные группы учащихся сажают разные виды семян, чтобы определить лучшую почву для прорастания семян. Я бы предложил использовать этот тип урока в классе после урока о семенах.
Навыки научного процесса:
Наблюдение, общение, измерение и вывод (AL Science, класс 3, концепция 21)
Отражение частей семени
Я чувствую, что урок под названием «Части семени» прошел очень хорошо, когда я провел его в классе миссис Хайнс. Дети были очень восприимчивы и с энтузиазмом относились к нашей деятельности. Я начал урок с учениками, сидящими на полу. Я спросил их, что они знают о семенах и как вода может изменить семена. Студенты предположили, что вода не повлияет на размер и цвет семени. Я провел их через дискуссию о семенах. Помогло то, что я заранее запланировал несколько вопросов, которые мог задать. В противном случае дискуссия не пошла бы так гладко, как прошла. На этом этапе мы прошлись по частям семени. У каждого ребенка есть журнал растений с семенами, которые нужно пометить. Учащиеся ответили на свои листы и вернулись к своим партам. Я считал это первой частью урока, и все прошло хорошо. Мои ученики прекрасно участвовали в обсуждении и были в восторге от того, что должно было произойти.
Когда ученики вернулись за свои парты, я раздал им рабочий лист «Внутри семени: часть 2» и велел им поместить его в свой дневник растений. Затем я раздал бумажное полотенце, влажную лимскую фасоль и сухую лимскую фасоль. Мне не помешала бы помощь при раздаче всех припасов. Процесс занял много времени, и студенты забеспокоились. Возможно, было бы полезно, если бы один ученик раздавал бумажные полотенца, а другой – сушеную лимскую фасоль. Влажные лимские бобы лучше всего раздавать учителю. Рабочий лист в журналах для учащихся содержит инструкции о том, что делать с мокрой и сухой фасолью. Они должны описывать, отслеживать, измерять и рисовать бобы на бумаге. Эта часть урока прошла гладко. Мои ученики с легкостью следовали инструкциям. После заполнения рабочего листа я попросила детей разорвать бобы и исследовать, что внутри. Они должны были попытаться понять работу частей семени. Я не знаю, ясно ли я изложил эти указания, потому что большинство студентов не искали части семени после того, как они вскрыли его. Мне пришлось ходить по комнате, спрашивая их по отдельности, видели ли они каждую часть. Это могло бы быть намного лучше, но в итоге все ученики увидели каждую часть семени.
В следующей части урока я вытащила детей на пол для коллоквиума. Я задавал вопросы, а студенты отвечали и обсуждали. Я хотел, чтобы ученики сами отвечали на свои вопросы, но обсуждение в основном вели учителя. У меня были заранее запланированные вопросы на случай, если это произойдет, но я действительно старался способствовать тому, чтобы они вели дискуссию. Мы рассказали об увиденном и работе каждой части семени. Я чувствую, что заранее спланированные вопросы помогли направить дискуссию в нужном мне направлении. Если бы мне пришлось проводить этот урок еще раз, я бы дал студентам больше времени, чтобы они ответили на свои вопросы и возглавили дискуссию.
Для оценочного упражнения детей отослали обратно к своим партам, попросили убрать кусочки фасоли/семян и дали пустое семя с этикетками, которые они должны были заполнить. какова была его функциональная работа. Эта страница была открыта в конце урока, чтобы я мог видеть, как много они узнали. Я думал о том, чтобы взяться за этот рабочий лист, но решил это сделать, чтобы дети знали, что мне интересно наблюдать за их прогрессом.